Качество каких электродов выше ок 93 95 или озана 2: Выбор сварочных электродов для стали: сравнение основных марок
Содержание
Электроды для сварки алюминия
Для сварки алюминиевых изделий, наплавки и устранения брака при отливке созданы специальные электроды. Применявшиеся ранее покрытые электроды не позволяли обеспечить нужный результат сварных работ.
Сегодняшние электроды для сварки алюминия обеспечивают высокое качество электродуговой сварки и применяются во многих сферах промышленности.
Электроды ОЗАНА-1 и ОЗАНА-2
Покрытые электроды ОЗАНА-1, начиная с 2001 года, широко используются для сварки и наплавки конструктивных элементов из технического алюминия.
Сварочные электроды ОЗАНА-2 применяются для устранения производственного брака при отливе и наплавки деталей из алюминиево-кремнистых сплавов. При этом сварка выполняется намного быстрее по сравнению со стальными электродами.
В процессе работы электрод используется без поступательно-возвратных движений и сжигается безостановочно. Данные электроды имеют стабильный режим горения, дают высококачественный шов, близкий по характеристикам к свариваемому металлу.
Данные электроды имеют стабильный режим горения, дают высококачественный шов, близкий по характеристикам к свариваемому металлу.
Актуальные цены 2023 года от поставщиков
Шлаковая корка после работ легко отделяется. Для предотвращения коррозии шлак счищают металлической щеткой, а сам шов промывают горячей водой.
Электроды ОК 96.10, ОК 96.20, ОК 96.50
Данные электроды применяются для сваривания литейных алюминиевых сплавов, включая силумины. Электроды ОК 96.10 имеют щелочно-солевое покрытие и алюминиево-кремниевый стержень. Режим сварки выбирается с учетом того, что скорость плавления ОК 96.10 втрое превышает скорость плавления стальных электродов.
Для проведения сварочных работ требуется предварительный нагрев от 150 °С до 300 °С. В электродах ОК 96.20 алюминиево-марганцевый стержень покрыт щелочно-солевым составом.
Изделия применяют для сварки прокатного свариваемого алюминия, в том числе алюминиево-марганцевых и алюминиево-магниевых конструкций для судостроительных, пивоваренных и молочно-перерабатывающих предприятий. Перед проведением сварки осуществляют нагрев до 150 – 300 °С.
Перед проведением сварки осуществляют нагрев до 150 – 300 °С.
Поскольку покрытие довольно восприимчиво к влаге, электроды необходимо хранить в сухих складах во влагозащитной упаковке. Если это условие не соблюдается, снижается качество сварочного шва.
Неплавящиеся электроды для сварки алюминия
Данный вид электродов изготавливается из вольфрама и применяется для проведения сварочных работ по алюминию в аргоновой среде. Вольфрамовые электроды имеют сечение в 2 -6 мм и выбираются в соответствии с толщиной свариваемого металла.
В качестве сырья для их производства используют как вольфрам в чистом виде, так и с различными добавками, улучшающими качество сварочного шва.
Видео: Сварка алюминия покрытыми электродами
Найти мастера
—
—
Читайте также:
Сварка полиэтиленовых труб
Сварочные электроды
Система аттестация сварочного производства накс
Повышение эффективности охлаждения газотурбинных камер сгорания
Современные технологии металлообработки
Пайка в домашних условиях
- Назад
- Вперёд
Электрод ОЗАНА-2 Ø3мм — 4мм
Выберите категорию
Все
Электроды сварочные
» Нержавейка и сварка высоколегированных сталей
»» Электрод АНЖР-1 Ø3мм — Ø5мм
»» Электрод АНЖР-2 Ø3мм. — Ø5мм.
— Ø5мм.
»» Электрод ЗИО-8 Ø2,0мм. — Ø5,0мм.
»» Электрод НЖ-13 Ø3мм — Ø5мм.
»» Электрод НИАТ-1 Ø2,0мм. — Ø5,0мм.
»» Электрод НИАТ-5 Ø2,0мм. — Ø5,0мм. / упаковка 5кг.
»» Электрод НИИ-48Г Ø3,0мм. — Ø5,0мм.
»» Электрод ОЗЛ-6 Ø3,0мм.- Ø4,0мм. ПЛАЗМА
»» Электрод ОЗЛ-6 Ø2,0мм — Ø5,0мм
»» Электрод ОЗЛ-8 Ø2,0мм — Ø5,0мм
»» Электрод ОЗЛ-9А Ø3,0мм. — Ø5,0мм.
— Ø5,0мм.
»» Электрод ОЗЛ-14А Ø3,0мм — Ø5,0мм
»» Электрод ОЗЛ-17У Ø3,0мм — Ø5,0мм.
»» Электрод ОЗЛ-19 Ø3,0мм — Ø5,0мм.
»» Электрод ОЗЛ-22 Ø3,0мм. — Ø5,0мм.
»» Электрод ОЗЛ-25Б Ø3,0мм. — Ø5,0мм.
»» Электрод ОЗЛ-25 Ø3,0мм. — Ø5,0мм.
»» Электрод ОЗЛ-36 Ø3,0мм. — Ø5,0мм.
»» Электрод УОНИ-13НЖ Ø3,0мм. — Ø5,0мм. 12Х13
»» Электрод ЦЛ-9 Ø3,0мм. — Ø5,0мм.
— Ø5,0мм.
»» Электрод ЦЛ-11 Ø3,0мм — Ø4,0мм Плазма
»» Электрод ЦЛ-11 Ø2,0мм. — Ø5,0мм.
»» Электрод ЦЛ-25/1 Ø3,0мм. — Ø5,0мм.
»» Электрод ЦТ-15 Ø2,0мм. — Ø5,0мм.
»» Электрод ЦТ-28 Ø3,0мм. — Ø4,0мм.
»» Электрод ЭА-395/9 Ø2,0мм. — Ø5,0мм.
»» Электрод ЭА-400/10Т Ø2,0мм. — Ø5,0мм.
»» Электрод ЭА-400/10У Ø2,0мм. — Ø5,0мм.
»» Электрод ЭА-898/21Б Ø3,0мм. — Ø5,0мм.
— Ø5,0мм.
»» Электрод ESAB ОК 61.30 Ø2,5мм. — Ø4,0мм
»» Электрод ESAB ОК 61.85 Ø2,5мм — Ø4,0мм
»» Электрод ESAB ОК 63.20 Ø2,5мм. — Ø3,2мм.
»» Электрод ESAB ОК 63.30 Ø2,5мм — Ø3,2ММ.
»» Электрод ESAB ОК 67.60 Ø2,5мм. — Ø3,2мм.
» Для чугуна
»» Электрод ESAB OK NiFe-Cl 2,5м — 4,0мм
»» Электрод МНЧ-2 Ø3мм -5мм
»» Электрод ОЗЧ-2 Ø3мм — 5мм
»» Электрод ОЗЧ-3 Ø3мм — 4мм
»» Электрод ОЗЧ-4 Ø3мм — 4мм
»» Электрод ОЗЧ-6 Ø3мм — 5мм
»» Электрод ЦЧ-4 Ø3мм — 5мм
»» Электрод ЦЧ-4 Монолит Ø3мм- 4мм
» Наплавка
»» Электрод ESAB OK NiFe-Cl-A
»» Электрод ВСН-6 Ø3мм — 5мм
»» Электрод НР-70 Ø3мм — 5мм
»» Электрод ОЗИ-6 Ø3мм — 5мм
»» Электрод ОЗН-6 Ø3мм — 5мм
»» Электрод ОЗШ-1 Ø3мм — 5мм
»» Электрод ОЗШ-6 Ø3мм — 5мм
»» Электрод ОМГ-Н Ø3мм — 5мм
»» Электрод Т-590 Ø4мм — 5мм
»» Электрод Т-620 Ø4мм — 5мм
»» Электрод ЦН-12М Ø3мм — 5мм
»» Электрод ЦН-6Л Ø3мм — 5мм
»» Электрод ЦНИИН-4 Ø3мм — 4мм
»» Электрод ЭН-60М Ø3мм — 5мм
»» Электрод УОНИ — 13НЖ 20Х13
» Цветные металлы
»» Электрод АНЦ/ОЗМ-3 Ø3мм — 5мм
»» Электрод КОМСОМОЛЕЦ-100 Ø3мм — 5мм
»» Электрод ОЗА-1 Ø3мм — 4мм
»» Электрод ОЗА-2 Ø3мм — 4мм
»» Электрод ОЗАНА-1 Ø3мм — 4мм
»» Электрод ОЗАНА-2 Ø3мм — 4мм
»» Электрод ОЗБ-2М Ø3мм — 5мм
»» Электроды ESAB OK 94. 25 Ø2,5мм — 3,2мм
25 Ø2,5мм — 3,2мм
» Теплоустойчивые стали
»» Электрод ESAB OK 74.70 Ø3,2мм — 4,0мм
»» Электрод ТМЛ-1У Ø3мм — 5мм
»» Электрод ТМЛ-3У Ø3мм — 5мм
»» Электрод ТМЛ-5 Ø3мм — 5мм
»» Электрод ТМУ-21У Ø3мм — 5мм
»» Электрод УОНИ-13/65 Ø3мм — 5мм
»» Электрод УОНИ-13/85 Ø3мм — 5мм
»» Электрод ЦЛ-17 Ø3мм — 5мм
»» Электрод ЦЛ-20 Ø3мм — 5мм
»» Электрод ЦЛ-39 Ø2,5мм
»» Электрод ЦУ-5 Ø2,5мм
» Углеродистые стали
»» Электрод Basic One Ø3мм — 5мм
»» Электрод ESAB OK 53. 70 ф2,5мм — 4мм
70 ф2,5мм — 4мм
»» Электрод ESAB ОЗС-12
»» Электрод ESAB ОК 48.00 Ø2,5мм — 4мм
»» Электрод ESAB ОК 48.04
»» Электрод LINCOLN ELECTRIC Omnia 46
»» Электрод LINCOLN ELECTRIC АНО-21
»» Электрод LINCOLN ELECTRIC МГМ-50К
»» Электрод LINCOLN ELECTRIC МР-3
»» Электрод LINCOLN ELECTRIC МР-3С
»» Электрод LINCOLN ELECTRIC ОЗС-12
»» Электрод LINCOLN ELECTRIC УОНИ-13/45
»» Электрод LINCOLN ELECTRIC УОНИ-13/55
»» Электрод Арсенал МР-3 ф3мм
»» Электрод Монолит АРМО МР-3
»» Электрод Монолит РЦ
»» Электрод МР-3 ф3мм ESAB
»» Электрод Спецэлектрод МР-3
»» Электрод Спецэлектрод МР-3С
»» Электрод Спецэлектрод УОНИ-13/55
»» Электрод УОНИ-13/55 Плазма
»» Электроды ESAB OK 46. 00
00
»» Электроды ESAB УОНИИ-13/55
»» Электрод LB-52U KOBELCO
»» Электрод УОНИ-13/55 ф3мм ОЛИВЕР (5кг)
»» Электрод МР-3 Премиум ОЛИВЕР
»» Электрод ОЗС-12 ОЛИВЕР Беларусь
Сварочная проволока (Катушка, моток, пруток)
» Высоколегированная проволока (Нержавейка) Мотки, Катушки, Прутки.
»» Проволока MIG ER-308LSi
»» Проволока MIG ER-309LSi
»» Проволока MIG ER-316LSi
»» Проволока MIG ER-321
»» Проволока MIG ER-347
»» Проволока Св-01Х19Н9
»» Проволока Св-04Х19Н9
»» Проволока Cв-04Х19Н11М3
»» Проволока Св-05Х20Н9ФБС
»» Проволока Св-06Х19Н9Т
»» Проволока Св-07Х19Н10Б
»» Проволока Св-07Х25Н13
»» Проволока Св-08Х19Н10Г2Б
»» Проволока Св-08Х20Н9Г7Т
»» Проволока Св-08Х21Н10Г6
»» Проволока Св-10Х16Н25АМ6
»» Проволока Св-10Х17Т
» Алюминиевая проволока и прутки для сварки
»» Проволока MIG ER-4043
»» Проволока MIG ER-5356
»» Пруток TIG ER-5183
» Низколегированная проволока
»» Lincoln Eleсtric МЦЕНСК
»» FARINA Китай
»» Св-08Г2С ESAB
»» OK Autrod 12. 50 ESAB
50 ESAB
»» OK Autrod 12.51 ESAB
»» Св-08А Мотки
»» Св-08А Прутки
»» Проволока порошковая Е71Т-1GS
Вольфрамовые неплавящиеся электроды
» Вольфрамовый электрод WС-20
» Вольфрамовый электрод WL-15
» Вольфрамовый электрод WL-20
» Вольфрамовый электрод WP
» Вольфрамовый электрод WТ-20
» Вольфрамовый электрод WY-20
» Вольфрамовый электрод WZ-8
КРУГИ отрезные, зачистные, лепестковые.
» «Лужский абразивный завод» — г.Луга
»» Круги отрезные
»» Круги зачистные (Обдирочные)
»» Круг лепестковый торцевой КЛТ 125х22
» «TIGARBO» — г. Каменск — Шахтинский
»» Отрезные круги «TIGARBO» — г. Каменск — Шахтинский
»» Зачистные круги «TIGARBO» — г. Каменск — Шахтинский
Гвозди строительные и винтовые
» Гвозди строительные ГОСТ 4028-63
» Гвозди винтовые ТУ 14-4-1161-2003
Газосварка
» Резаки Газовые
»» Резаки пропановые
»» Резаки ацетиленовые
»» Резаки универсальные
» Горелки газовые (Ацетилен, Пропан)
Производитель
ВсеESABESAB (СВЭЛ)ESAB (СВЭЛ) РоссияESAB / Китай / ИндияFARINA КитайGoldSparkLINCOLN ELECTRIC / МежгосМетиз МценскLinkoln ElectricАрсенал (Белоруссия)ВЕКТОРДОН РоссияДОНМЕТДОНМЕТ (Украина)ИндияКитайЛосиноостровский Электродный завод (ЛЭЗ)Лужский абразивный заводНОРД (Россия)НПО СпецЭлектрод (г. Волгодонск), ПКФ ЯрЭМП (г. Ярославль)НПО Спецэлектрод ВолгодонскОливер БеларусьПлазма TM Monolith БелоруссияПлазма ТМ MonolithРОАРРоссияСеверсталь-МетизСЗСЭ БелоруссияСпецЭлектрод МоскваСудиславский Завод СМ (СЗСМ)ТIGARBO г. Каменск-ШахтинскийЭСАБ — ТюменьЯпонияЯпония/ИндияЯрЭМП (Ярославль)
Волгодонск), ПКФ ЯрЭМП (г. Ярославль)НПО Спецэлектрод ВолгодонскОливер БеларусьПлазма TM Monolith БелоруссияПлазма ТМ MonolithРОАРРоссияСеверсталь-МетизСЗСЭ БелоруссияСпецЭлектрод МоскваСудиславский Завод СМ (СЗСМ)ТIGARBO г. Каменск-ШахтинскийЭСАБ — ТюменьЯпонияЯпония/ИндияЯрЭМП (Ярославль)
Гомозиготная экспрессия мутантного ELOVL4 приводит к судорогам и смерти в новой животной модели дефицита жирных кислот с очень длинной цепью , MacDonald IM, Sieving PA, Figueroa DJ, Austin CP, Gould RJ, Ayyagari R, Petrukhin K. Делеция 5 п.н. в ELOVL4 связана с двумя родственными формами аутосомно-доминантной макулярной дистрофии. Нат Жене. 2001;27(1):89–93. дои: 10.1038/83817. [PubMed] [CrossRef] [Академия Google]
2. Bernstein PS, Tammur J, Singh N, Hutchinson A, Dixon M, Pappas CM, Zabriskie NA, Zhang K, Petrukhin K, Leppert M, Allikmets R. Разнообразный фенотип макулярной дистрофии, вызванный новой сложной мутацией в гене ELOVL4. Invest Ophthalmol Vis Sci. 2001;42(13):3331–3336. [PubMed] [Google Scholar]
3. Эдвардс А.О., Доносо Л.А., Риттер Р., 3-й Новый ген аутосомно-доминантной макулярной дистрофии, напоминающей Штаргардта, с гомологией белков семейства SUR4. Invest Ophthalmol Vis Sci. 2001;42(11):2652–2663. [PubMed] [Академия Google]
Эдвардс А.О., Доносо Л.А., Риттер Р., 3-й Новый ген аутосомно-доминантной макулярной дистрофии, напоминающей Штаргардта, с гомологией белков семейства SUR4. Invest Ophthalmol Vis Sci. 2001;42(11):2652–2663. [PubMed] [Академия Google]
4. Agbaga MP, Brush RS, Mandal MN, Henry K, Elliott MH, Anderson RE. Роль белка макулярной дистрофии Штаргардта-3 (ELOVL4) в биосинтезе жирных кислот с очень длинной цепью. Proc Natl Acad Sci USA. 2008;105(35):12843–12848. doi: 10.1073/pnas.0802607105. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
5. Logan S, Agbaga MP, Chan MD, Brush RS, Anderson RE. Микроокружение эндоплазматического ретикулума и консервативные гистидины регулируют активность элонгазы жирных кислот ELOVL4. J липидный рез. 2014;55(4):698–708. doi: 10.1194/jlr.M045443. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
6. Uchida Y, Holleran WM (2008) Омега-O-ацилцерамид, липид, необходимый для выживания млекопитающих. J Dermatol Sci 51 (2): 77–87. 10.1016/j.jdermsci.2008.01.002 [PubMed]
10.1016/j.jdermsci.2008.01.002 [PubMed]
7. Cameron DJ, Tong Z, Yang Z, Kaminoh J, Kamiyah S, Chen H, Zeng J, Chen Y, Luo L, Zhang K. Существенная роль Elovl4 в синтезе очень длинноцепочечных жирных кислот, барьерной функции кожной проницаемости и выживаемости новорожденных. Int J Biol Sci. 2007;3(2):111–119.. doi: 10.7150/ijbs.3.111. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Li W, Sandhoff R, Kono M, Zerfas P, Hoffmann V, Ding BC, Proia RL, Deng CX. Истощение церамидов жирными кислотами с очень длинной цепью вызывает нарушение барьерной функции кожной проницаемости и неонатальную летальность у мышей с дефицитом ELOVL4. Int J Biol Sci. 2007;3(2):120–128. doi: 10.7150/ijbs.3.120. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. McMahon A, Butovich IA, Mata NL, Klein M, Ritter R, 3rd, Richardson J, Birch DG, Edwards AO, Kedzierski W. Патология сетчатки и дефект кожного барьера у мышей, несущих мутацию болезни Штаргардта-3 в элонгазе очень длинноцепочечных жирных кислот-4 . Мол Вис. 2007; 13: 258–272. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мол Вис. 2007; 13: 258–272. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Vasireddy V, Uchida Y, Salem N, Jr, Kim SY, Mandal MN, Reddy GB, Bodepudi R, Alderson NL, Brown JC, Hama H, Dlugosz A, Elias PM, Holleran WM, Ayyagari R. Потеря функционального ELOVL4 истощает жирные кислоты с очень длинной цепью (> или = C28) и уникальный o мега-O-ацилцерамиды в коже, приводящие к неонатальной смерти. Хум Мол Жене. 2007;16(5):471–482. doi: 10.1093/hmg/ddl480. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. McMahon A, Lu H, Butovich IA. Роль ELOVL4 в мейбомиевой железе мыши и биологии клеток себоцитов. Invest Ophthalmol Vis Sci. 2014;55(5):2832–2840. doi: 10.1167/iovs.13-13335. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Brush RS, Tran JT, Henry KR, McClellan ME, Elliott MH, Mandal MN. Сфинголипиды сетчатки и их виды, содержащие жирные кислоты с очень длинной цепью. Invest Ophthalmol Vis Sci. 2010;51(9):4422–4431. doi: 10.1167/iovs. 09-5134. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
09-5134. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Авелдано М.И. Новая группа полиеновых жирных кислот с очень длинной цепью в диполиненасыщенных фосфатидилхолинах сетчатки позвоночных. Дж. Биол. Хим. 1987;262(3):1172–1179. [PubMed] [Академия Google]
14. Poulos A, Johnson DW, Beckman K, White IG, Easton C. Возникновение необычных молекулярных видов сфингомиелина, содержащих 28-34-углеродные полиеновые жирные кислоты, в сперматозоидах барана. Биохим Дж. 1987;248(3):961–964. doi: 10.1042/bj2480961. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15. Zanetti SR, Maldonado EN, Aveldano MI. Доксорубицин воздействует на липиды яичек длинноцепочечными (С18-С22) и очень длинноцепочечными (С24-С32) полиненасыщенными жирными кислотами. Рак рез. 2007;67(14):6973–6980. doi: 10.1158/0008-5472.CAN-07-0376. [PubMed] [CrossRef] [Google Scholar]
16. Авелдано М.И. Длинные и очень длинные полиненасыщенные жирные кислоты сетчатки и сперматозоидов: полный набор полиеновых жирных кислот. Adv Exp Med Biol. 1992; 318: 231–242. doi: 10.1007/978-1-4615-3426-6_19. [PubMed] [CrossRef] [Google Scholar]
Adv Exp Med Biol. 1992; 318: 231–242. doi: 10.1007/978-1-4615-3426-6_19. [PubMed] [CrossRef] [Google Scholar]
17. Одзаки К., Дои Х., Мицуи Дж., Сато Н., Иикуни Ю., Мадзима Т., Ямане К., Ириока Т., Ишиура Х., Дои К., Моришита С., Хигаси М., Секигути Т., Кояма К., Уэда Н., Миура Й., Миятакэ С., Мацумото Н., Yokota T, Tanaka F, Tsuji S, Mizusawa H, Ishikawa K. Новая мутация в ELOVL4, приводящая к спиноцеребеллярной атаксии (SCA) с признаком горячей поперечной булочки, но без эритрокератодермии: расширенный спектр SCA34. ДЖАМА Нейрол. 2015;72(7):797–805. doi: 10.1001/jamaneurol.2015.0610. [PubMed] [CrossRef] [Google Scholar]
18. Mir H, Raza SI, Touseef M, Memon MM, Khan MN, Jaffar S, Ahmad W. Новая рецессивная мутация в гене ELOVL4 вызывает нейро-ихтиотическое расстройство с переменной экспрессивностью. БМС Мед Жене. 2014;15(1):25. дои: 10.1186/1471-2350-15-25. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
19. Cadieux-Dion M, Turcotte-Gauthier M, Noreau A, Martin C, Meloche C, Gravel M, Drouin CA, Rouleau GA, Nguyen DK, Cossette P. Расширение клинического фенотипа, связанного с мутацией ELOVL4: исследование большого франко-канадского семья с аутосомно-доминантной спиноцеребеллярной атаксией и эритрокератодермией. ДЖАМА Нейрол. 2014;71(4):470–475. doi: 10.1001/jamaneurol.2013.6337. [PubMed] [CrossRef] [Академия Google]
Расширение клинического фенотипа, связанного с мутацией ELOVL4: исследование большого франко-канадского семья с аутосомно-доминантной спиноцеребеллярной атаксией и эритрокератодермией. ДЖАМА Нейрол. 2014;71(4):470–475. doi: 10.1001/jamaneurol.2013.6337. [PubMed] [CrossRef] [Академия Google]
20. Bourassa CV, Raskin S, Serafini S, Teive HA, Dion PA, Rouleau GA. Новая мутация ELOVL4 при спиноцеребеллярной атаксии с эритрокератодермией. ДЖАМА Нейрол. 2015;72(8):942–943. doi: 10.1001/jamaneurol.2015.0888. [PubMed] [CrossRef] [Google Scholar]
21. Aldahmesh MA, Mohamed JY, Alkuraya HS, Verma IC, Puri RD, Alaiya AA, Rizzo WB, Alkuraya FS. Рецессивные мутации в ELOVL4 вызывают ихтиоз, умственную отсталость и спастическую квадриплегию. Am J Hum Genet. 2011;89(6): 745–750. doi: 10.1016/j.ajhg.2011.10.011. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22. Agbaga MP. Различные мутации в ELOVL4 влияют на биосинтез жирных кислот с очень длинной цепью, вызывая различные неврологические расстройства у людей. Adv Exp Med Biol. 2016; 854: 129–135. doi: 10.1007/978-3-319-17121-0_18. [PubMed] [CrossRef] [Google Scholar]
Adv Exp Med Biol. 2016; 854: 129–135. doi: 10.1007/978-3-319-17121-0_18. [PubMed] [CrossRef] [Google Scholar]
23. Logan S, Agbaga MP, Chan MD, Kabir N, Mandal NA, Brush RS, Anderson RE. Расшифровка активности мутанта ELOVL4 при аутосомно-доминантной макулярной дистрофии Штаргардта. Proc Natl Acad Sci U S A. 2013;110(14):5446–5451. doi: 10.1073/pnas.1217251110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Шерри Д.М., Хопиавуори Б.Р., Стайлз М.А., Рахман Н.С., Озан К.Г., Дик Ф., Агбага М.П., Андерсон Р.Е. (2017)Распределение ELOVL4 в мозге развивающихся и взрослых мышей. Фронт Нейроанат 11(38). 10.3389/fnana.2017.00038 [бесплатная статья PMC] [PubMed]
25. Mandal MN, Ambasudhan R, Wong PW, Gage PJ, Sieving PA, Ayyagari R. Характеристика мышиного ортолога ELOVL4: геномная организация и пространственно-временная экспрессия. Геномика. 2004;83(4):626–635. doi: 10.1016/j.ygeno.2003.09.020. [PubMed] [CrossRef] [Академия Google]
26. McMahon A, Butovich IA, Kedzierski W. Эпидермальная экспрессия трансгена Elovl4 спасает неонатальную смертность гомозиготных мышей с болезнью Штаргардта-3. J липидный рез. 2011;52(6):1128–1138. doi: 10.1194/jlr.M014415. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
Эпидермальная экспрессия трансгена Elovl4 спасает неонатальную смертность гомозиготных мышей с болезнью Штаргардта-3. J липидный рез. 2011;52(6):1128–1138. doi: 10.1194/jlr.M014415. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
27. Вассар Р., Розенберг М., Росс С., Тайнер А., Фукс Э. Тканеспецифичная и специфичная для дифференцировки экспрессия гена кератина человека K14 у трансгенных мышей. Proc Natl Acad Sci U S A. 1989;86(5):1563–1567. doi: 10.1073/pnas.86.5.1563. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Vassar R, Fuchs E. Трансгенные мыши позволяют по-новому взглянуть на роль TGF-альфа во время эпидермального развития и дифференцировки. Гены Дев. 1991;5(5):714–727. doi: 10.1101/gad.5.5.714. [PubMed] [CrossRef] [Google Scholar]
29. Ромейн Х. Дж., ван Уум Дж. Ф., Бридейк И., Эммеринг Дж., Раду И., Пул CW. Двойное иммуномечение нейропептидов в гипоталамусе человека по результатам анализа с помощью конфокальной лазерной сканирующей флуоресцентной микроскопии. J Гистохим Цитохим. 1999;47(2):229–236. дои: 10.1177/002215549904700211. [PubMed] [CrossRef] [Google Scholar]
J Гистохим Цитохим. 1999;47(2):229–236. дои: 10.1177/002215549904700211. [PubMed] [CrossRef] [Google Scholar]
30. Ricciuti A, De Remigis A, Landek-Salgado MA, De Vincentiis L, Guaraldi F, Lupi I, Iwama S, Wand GS, Salvatori R, Caturegli P. Обнаружение гипофизарных антител с помощью иммунофлуоресценции: подход и приводит к заболеваниям гипофиза. J Clin Endocrinol Metab. 2014; 99(5):1758–1766. doi: 10.1210/jc.2014-1049. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Bennett LD, Brush RS, Chan M, Lydic TA, Reese K, Reid GE, Busik JV, Elliott MH, Anderson RE. Влияние восстановленных VLC-PUFA сетчатки на фоторецепторы палочек и колбочек. Invest Ophthalmol Vis Sci. 2014;55(5):3150–3157. doi: 10.1167/iovs.14-13995. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Bennett LD, Hopiavuori BR, Brush RS, Chan M, Van Hook MJ, Thoreson WB, Anderson RE. Исследование терминалей фоторецепторов с дефицитом VLC-PUFA. Invest Ophthalmol Vis Sci. 2014;55(7):4063–4072. doi: 10.1167/iovs.14-13997. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
doi: 10.1167/iovs.14-13997. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Mullen RJ, Buck CR, Smith AM. NeuN, специфический для нейронов ядерный белок позвоночных. Разработка. 1992;116(1):201–211. [PubMed] [Академия Google]
34. Wolf HK, Buslei R, Schmidt-Kastner R, Schmidt-Kastner PK, Pietsch T, Wiestler OD, Blumcke I. NeuN: полезный нейрональный маркер для диагностической гистопатологии. J Гистохим Цитохим. 1996;44(10):1167–1171. doi: 10.1177/44.10.8813082. [PubMed] [CrossRef] [Google Scholar]
35. Chang YC, Gottlieb DI. Характеристика белков, очищенных моноклональными антителами к декарбоксилазе глутаминовой кислоты. Дж. Нейроски. 1988;8(6):2123–2130. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Бакли К., Келли Р.Б. Идентификация трансмембранного гликопротеина, специфичного для секреторных пузырьков нервных и эндокринных клеток. Джей Селл Биол. 1985; 100(4):1284–1294. doi: 10.1083/jcb.100.4.1284. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. VanGuilder HD, Brucklacher RM, Patel K, Ellis RW, Freeman WM, Barber AJ. Диабет подавляет пресинаптические белки и снижает фосфорилирование базального синапсина I в сетчатке крыс. Евр Джей Нейроски. 2008;28(1):1–11. дои: 10.1111/j.1460-9568.2008.06322.х. [PubMed] [CrossRef] [Google Scholar]
VanGuilder HD, Brucklacher RM, Patel K, Ellis RW, Freeman WM, Barber AJ. Диабет подавляет пресинаптические белки и снижает фосфорилирование базального синапсина I в сетчатке крыс. Евр Джей Нейроски. 2008;28(1):1–11. дои: 10.1111/j.1460-9568.2008.06322.х. [PubMed] [CrossRef] [Google Scholar]
38. VanGuilder HD, Yan H, Farley JA, Sonntag WE, Freeman WM. Старение изменяет экспрессию белков, регулирующих нейротрансмиссию, в синаптопротеоме гиппокампа. Дж. Нейрохим. 2010;113(6):1577–1588. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Hopiavuori BR, Masser DR, Wilkerson JL, Brush RS, Mandal NA, Anderson RE, Freeman WM. Выделение синаптических мембран нейронов центрифугированием в градиенте сахарозы. Методы Мол Биол. 2017;1609: 33–41. doi: 10.1007/978-1-4939-6996-8_4. [PubMed] [CrossRef] [Google Scholar]
40. Bligh EG, Dyer WJ. Экспресс-метод экстракции и очистки общих липидов. Может J Biochem Physiol. 1959; 37 (8): 911–917. doi: 10.1139/o59-099. [PubMed] [CrossRef] [Google Scholar]
41. Li F, Marchette LD, Brush RS, Elliott MH, Le YZ, Henry KA, Anderson AG, Zhao C, Sun X, Zhang K, Anderson RE. DHA не защищает трансгенных мышей ELOVL4 от дегенерации сетчатки. Мол Вис. 2009; 15:1185–1193. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Li F, Marchette LD, Brush RS, Elliott MH, Le YZ, Henry KA, Anderson AG, Zhao C, Sun X, Zhang K, Anderson RE. DHA не защищает трансгенных мышей ELOVL4 от дегенерации сетчатки. Мол Вис. 2009; 15:1185–1193. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Лэрд Н.М., Уэр Дж.Х. Модели случайных эффектов для продольных данных. Биометрия. 1982;38(4):963–974. дои: 10.2307/2529876. [PubMed] [CrossRef] [Google Scholar]
43. Pinheiro JC, Bates DM. Модели со смешанными эффектами в S и S-PLUS. Нью-Йорк: Статистика и вычисления. Спрингер; 2000. [Google Scholar]
44. Venables WN, Ripley BD, Venables WN. Современная прикладная статистика с С. 4. Нью-Йорк: Статистика и вычислительная техника. Спрингер; 2002. [Google Scholar]
45. Вуд С.Н. Обобщенные аддитивные модели: введение с R. Texts в статистическую науку. Бока-Ратон: Чепмен и Холл/CRC; 2006. [Google Академия]
46. Бетц В.Дж., Бьюик Г.С. Оптический анализ рециркуляции синаптических пузырьков в нервно-мышечном соединении лягушки. Наука. 1992;255(5041):200–203. doi: 10.1126/science.1553547. [PubMed] [CrossRef] [Google Scholar]
Наука. 1992;255(5041):200–203. doi: 10.1126/science.1553547. [PubMed] [CrossRef] [Google Scholar]
47. Gaffield MA, Betz WJ. Визуализация экзоцитоза и эндоцитоза синаптических пузырьков с помощью красителей FM. Нат Проток. 2006;1(6):2916–2921. doi: 10.1038/nprot.2006.476. [PubMed] [CrossRef] [Google Scholar]
48. Deák F, Xu Y, Chang WP, Dulubova I, Khvotchev M, Liu X, Südhof TC, Rizo J. Связывание Munc18-1 с нейронным комплексом SNARE контролирует заправку синаптических пузырьков. Джей Селл Биол. 2009 г.;184(5):751–764. doi: 10.1083/jcb.200812026. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Deák F, Shin OH, Kavalali ET, Südhof TC. Структурные детерминанты функции синаптобревина 2 при слиянии синаптических пузырьков. Дж. Нейроски. 2006;26(25):6668–6676. doi: 10.1523/JNEUROSCI.5272-05.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Deák F, Schoch S, Liu X, Südhof TC, Kavalali ET. Синаптобревин необходим для быстрого эндоцитоза синаптических пузырьков. Nat Cell Biol. 2004;6(11):1102–1108. doi: 10.1038/ncb1185. [PubMed] [CrossRef] [Академия Google]
Nat Cell Biol. 2004;6(11):1102–1108. doi: 10.1038/ncb1185. [PubMed] [CrossRef] [Академия Google]
51. Деак Ф., Шин О.Х., Танг Дж., Хэнсон П., Убах Дж., Ян Р., Ризо Дж., Кавалали Э.Т., Зюдхоф Т.К. Рабфилин регулирует SNARE-зависимую перезапуск синаптических везикул для слияния. EMBO J. 2006; 25 (12): 2856–2866. doi: 10.1038/sj.emboj.7601165. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Навидхамиди М., Гасеми М., Мехранфард Н. Связанные с эпилепсией изменения возбудимости гиппокампа. Преподобный Нейроски. 2017;28(3):307–334. doi: 10.1515/revneuro-2016-0059. [PubMed] [CrossRef] [Академия Google]
53. Soussi R, Boulland JL, Bassot E, Bras H, Coulon P, Chaudhry FA, Storm-Mathisen J, Ferhat L, Escclaez M. Реорганизация надмаммилярно-гиппокампальных путей в крысиной пилокарпиновой модели эпилепсии височной доли: свидетельство прорастания терминала аксона. Структура мозга Функц. 2015;220(4):2449–2468. doi: 10.1007/s00429-014-0800-2. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
54. Deak F, Liu X, Khvotchev M, Li G, Kavalali ET, Sugita S, Sudhof TC. Альфа-латротоксин стимулирует новый путь Са2+-зависимого синаптического экзоцитоза, независимый от классического механизма синаптического слияния. Дж. Нейроски. 2009 г.;29(27):8639–8648. doi: 10.1523/JNEUROSCI.0898-09.2009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Deak F, Liu X, Khvotchev M, Li G, Kavalali ET, Sugita S, Sudhof TC. Альфа-латротоксин стимулирует новый путь Са2+-зависимого синаптического экзоцитоза, независимый от классического механизма синаптического слияния. Дж. Нейроски. 2009 г.;29(27):8639–8648. doi: 10.1523/JNEUROSCI.0898-09.2009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
55. Belanger M, Allaman I, Magistretti PJ. Энергетический метаболизм мозга: внимание на метаболическом сотрудничестве астроцитов и нейронов. Клеточный метаб. 2011;14(6):724–738. doi: 10.1016/j.cmet.2011.08.016. [PubMed] [CrossRef] [Google Scholar]
56. Кэш С., Юсте Р. Линейное суммирование возбуждающих входов пирамидными нейронами СА1. Нейрон. 1999;22(2):383–394. дои: 10.1016/S0896-6273(00)81098-3. [PubMed] [CrossRef] [Google Scholar]
57. Hao J, Wang XD, Dan Y, Poo MM, Zhang XH. Арифметическое правило пространственного суммирования возбуждающих и тормозных входов в пирамидных нейронах. Proc Natl Acad Sci U S A. 2009;106(51):21906–21911. doi: 10.1073/pnas.0912022106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
doi: 10.1073/pnas.0912022106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
58. Poirazi P, Brannon T, Mel BW. Пирамидальный нейрон как двухслойная нейронная сеть. Нейрон. 2003;37(6):989–999. doi: 10.1016/S0896-6273(03)00149-1. [PubMed] [CrossRef] [Академия Google]
59. Поирази П., Брэннон Т., Мел Б.В. Арифметика подпороговой синаптической суммации в модельной пирамидной клетке СА1. Нейрон. 2003;37(6):977–987. doi: 10.1016/S0896-6273(03)00148-X. [PubMed] [CrossRef] [Google Scholar]
60. Rohrbough J, Rushton E, Palanker L, Woodruff E, Matties HJ, Acharya U, Acharya JK, Broadie K. Церамидаза регулирует экзоцитоз и транспортировку синаптических пузырьков. Дж. Нейроски. 2004;24(36):7789–7803. doi: 10.1523/JNEUROSCI.1146-04.2004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
61. Эль Баваб С., Уста Дж., Родди П., Шульц З.М., Белавска А., Ханнун Ю.А. Субстратная специфичность церамидазы головного мозга крысы. J липидный рез. 2002;43(1):141–148. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
62. Ben Gedalya T, Loeb V, Israel E, Altschuler Y, Selkoe DJ, Sharon R. Альфа-синуклеин и полиненасыщенные жирные кислоты способствуют клатрин-опосредованному эндоцитозу и рециркуляции синаптических пузырьков. Трафик. 2009;10(2):218–234. doi: 10.1111/j.1600-0854.2008.00853.x. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. Teixeira G, Vieira LB, Gomez MV, Guatimosim C. Холестерин как ключевой игрок в балансе вызванного и спонтанного высвобождения глутамата в синаптосомах коры головного мозга крысы. Нейрохим Интерн. 2012;61(7):1151–1159. doi: 10.1016/j.neuint.2012.08.008. [PubMed] [CrossRef] [Google Scholar]
64. Линетти А., Фратанджели А., Таверна Э., Валнегри П., Франколини М., Каппелло В., Маттеоли М., Пассафаро М., Роза П. Снижение уровня холестерина нарушает экзоцитоз синаптических пузырьков. Дж. Клеточные науки. 2010; 123 (часть 4): 595–605. doi: 10.1242/jcs.060681. [PubMed] [CrossRef] [Google Scholar]
65. Kato K, Clark GD, Bazan NG, Zorumski CF. Фактор активации тромбоцитов как потенциальный ретроградный мессенджер в долгосрочной потенциации гиппокампа CA1. Природа. 1994;367(6459):175–179. дои: 10.1038/367175a0. [PubMed] [CrossRef] [Google Scholar]
Kato K, Clark GD, Bazan NG, Zorumski CF. Фактор активации тромбоцитов как потенциальный ретроградный мессенджер в долгосрочной потенциации гиппокампа CA1. Природа. 1994;367(6459):175–179. дои: 10.1038/367175a0. [PubMed] [CrossRef] [Google Scholar]
66. Izquierdo I, Fin C, Schmitz PK, Da Silva RC, Jerusalinsky D, Quillfeldt JA, Ferreira MB, Medina JH, Bazan NG. Улучшение памяти путем интрагиппокампального, интраамигдалального или интраэнторинального введения фактора активации тромбоцитов, измеренного в задаче на избегание торможения. Proc Natl Acad Sci U S A. 1995;92(11):5047–5051. doi: 10.1073/pnas.92.11.5047. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
67. Clark GD, Happel LT, Zorumski CF, Bazan NG. Усиление возбуждающей синаптической передачи в гиппокампе фактором активации тромбоцитов. Нейрон. 1992;9(6):1211–1216. doi: 10.1016/0896-6273(92)
-R. [PubMed] [CrossRef] [Google Scholar]
68. Chen C, Bazan NG. Липидная сигнализация: сон, синаптическая пластичность и нейропротекция. Простагландины Другие липиды Медиат. 2005;77(1–4):65–76. doi: 10.1016/j.prostaglandins.2005.07.001. [PubMed] [CrossRef] [Академия Google]
Преимущества независимых устройств DBS, управляемых контактным током
Профессор и заведующий кафедрой неврологии Миннесотского университета рассказал о недавних результатах исследования устройства Boston Scientific, управляемого несколькими независимыми контактными токами для глубокой стимуляции мозга.
Jerrold L. Vitek, MD, PhD, профессор и заведующий кафедрой неврологии Университета Миннесоты
Jerrold L. Vitek, MD, PhD
Контролируемая (MICC) система DBS от Boston Scientific безопасна и эффективна при лечении двигательных симптомов болезни Паркинсона.
В исследовании, проведенном Джерролдом Л. Витеком, доктором медицинских наук, профессором и заведующим кафедрой неврологии Миннесотского университета, и его коллегами, оценивалась клиническая эффективность и безопасность системы Vercise DBS у 191 пациента с болезнью Паркинсона и двигательными симптомами. Исследование, получившее название INTREPID (NCT01839396), показало, что от исходного уровня после имплантации до 3 месяцев после рандомизации разница в среднем изменении возрастала во времени без неприятных дискинезий между активными (n = 121) и контрольными (n = 39)) составил 3,03 часа (стандартное отклонение [SD], 4,52; 95% ДИ, 1,3–4,7; P <0,0001).
Исследование, получившее название INTREPID (NCT01839396), показало, что от исходного уровня после имплантации до 3 месяцев после рандомизации разница в среднем изменении возрастала во времени без неприятных дискинезий между активными (n = 121) и контрольными (n = 39)) составил 3,03 часа (стандартное отклонение [SD], 4,52; 95% ДИ, 1,3–4,7; P <0,0001).
Чтобы узнать больше о самом устройстве и о том, как эта технология MICC может предложить врачам больше, чем стандартная технология DBS, NeurologyLive обратился к Витеку, который поделился своим видением и опытом работы с устройством.
NeurologyLive : Что побудило к исследованию этого конкретного устройства?
Джеррольд Л. Витек, доктор медицины, доктор философии: Ну, я думаю, что производители устройств в целом, если они хотят получить одобрение FDA, чтобы они могли коммерциализировать свои устройства, они должны пройти испытание в рамках одобрения FDA. Им нужно показать, что это безопасно и эффективно, даже если они внесут незначительную модификацию своего устройства по сравнению с другим устройством, которое уже есть на рынке. На данный момент 3 компании, производящие устройства, имеют одобрение FDA, и я знаю обо всем, что мне нужно было провести клиническое испытание.
Им нужно показать, что это безопасно и эффективно, даже если они внесут незначительную модификацию своего устройства по сравнению с другим устройством, которое уже есть на рынке. На данный момент 3 компании, производящие устройства, имеют одобрение FDA, и я знаю обо всем, что мне нужно было провести клиническое испытание.
Medtronic начала с первого 1, Abbott сделала еще один, а теперь Boston Scientific сделала этот. У них действительно была уникальная система в том смысле, что для каждого из контактов, находящихся на проводе, было несколько источников тока, в то время как каждое другое устройство было единственным источником тока.
Имея опыт использования, чем он для вас отличается от используемых в настоящее время устройств DBS?
Что ж, мы всегда спорили об этом, когда много лет назад впервые получили эти устройства от Medtronic, первой группы, которая выпустила их, они были управляемы напряжением, а не током, и это был один источник. Если сопротивление вокруг контакта, который мы фактически активируем и инициируем ток от импеданса, изменяется, то величина протекающего тока изменяется. Оно может варьироваться в зависимости от сопротивления вокруг электрода. Некоторые люди показали, как это может меняться со временем, обычно самые большие изменения происходят в первые пару часов, но иногда вы можете увидеть эти изменения в течение более длительного периода. Допустим, я устанавливаю ваш стимулятор на определенный уровень, и это дает вам хороший контроль, но внезапно сопротивление вокруг электрода со временем увеличивается, тогда вы получаете меньший ток, и тогда ваши симптомы могут начать возвращаться. А потом ты вернешься и увидишь меня, и тогда нам нужно будет снова все отрегулировать.
Оно может варьироваться в зависимости от сопротивления вокруг электрода. Некоторые люди показали, как это может меняться со временем, обычно самые большие изменения происходят в первые пару часов, но иногда вы можете увидеть эти изменения в течение более длительного периода. Допустим, я устанавливаю ваш стимулятор на определенный уровень, и это дает вам хороший контроль, но внезапно сопротивление вокруг электрода со временем увеличивается, тогда вы получаете меньший ток, и тогда ваши симптомы могут начать возвращаться. А потом ты вернешься и увидишь меня, и тогда нам нужно будет снова все отрегулировать.
Но с устройством постоянного тока хорошо то, что вы можете просто установить ток, и вы получите такой большой ток независимо от сопротивления. Теперь есть и другие устройства постоянного тока. Но это было одно из первых устройств, которое действительно это сделало. И тот факт, что они могут контролировать количество тока, проходящего через каждый контакт, а в этом конкретном устройстве у них было 8 контактов. Также есть кое-что новое, называемое сегментированным выводом, который на самом деле имеет 8 контактов, но он настроен так, что это 1 круговой контакт, например, металлический круг, а затем 3 сегмента на следующем, 3 сегмента на следующем, а затем 1 нижний — называется 1-3-3-1. Сейчас у Бостона есть это устройство, но испытание было не таким. В этом испытании было 8 контактов, один из которых был наложен поверх другого. И опять же, преимущество здесь заключалось в том, что мы могли пропускать столько же или мало тока через каждый контакт.
Также есть кое-что новое, называемое сегментированным выводом, который на самом деле имеет 8 контактов, но он настроен так, что это 1 круговой контакт, например, металлический круг, а затем 3 сегмента на следующем, 3 сегмента на следующем, а затем 1 нижний — называется 1-3-3-1. Сейчас у Бостона есть это устройство, но испытание было не таким. В этом испытании было 8 контактов, один из которых был наложен поверх другого. И опять же, преимущество здесь заключалось в том, что мы могли пропускать столько же или мало тока через каждый контакт.
Идея заключается в том, что если вы стимулируете через один контакт, вы получите шар тока. Но с этим вы можете сделать мяч таким большим или маленьким, как вы хотите. Так что это было хорошо, потому что у вас может быть контакт, который действительно находится рядом с областью, в которой вам не нужен очень большой ток мяча, вам нужно немного, потому что вы можете его настроить. Затем вы можете подать немного тока и получить маленький шарик, затем чуть выше контакт может быть немного больше, и, может быть, тот, что выше, вы можете сделать немного меньше, или вы можете объединить два, верно? У вас есть много вариантов с точки зрения формирования текущего поля. Тип структур, в которые мы их помещаем, они не похожи на большие круги, они имеют геометрическую форму. И затем, где бы ни закончилось ваше упреждение, вы можете захотеть сделать мяч меньшего размера или мяч большего размера в разных местах. Это дает вам такую гибкость.
Тип структур, в которые мы их помещаем, они не похожи на большие круги, они имеют геометрическую форму. И затем, где бы ни закончилось ваше упреждение, вы можете захотеть сделать мяч меньшего размера или мяч большего размера в разных местах. Это дает вам такую гибкость.
Теперь, когда у них есть этот новый 1-3-3-1, у Boston Scientific и Abbot есть это преимущество, но теперь у Boston есть возможность направить ток через сегмент в одном направлении, а не подавать его в 1 из этих 8 контактов, что будет просто шаром, и теперь они могут перемещать его в разных направлениях. В этом исследовании использовалось примерно 8 контактов, один поверх другого, и вы можете контролировать, какой ток проходит через каждый контакт. В этом было преимущество.
Что клиническое сообщество должно вынести из этих результатов?
Знаете, было много исследований, если бы вы заглянули в PubMed и использовали глубокую стимуляцию мозга, вы бы удивились. Произошел экспоненциальный рост числа исследований.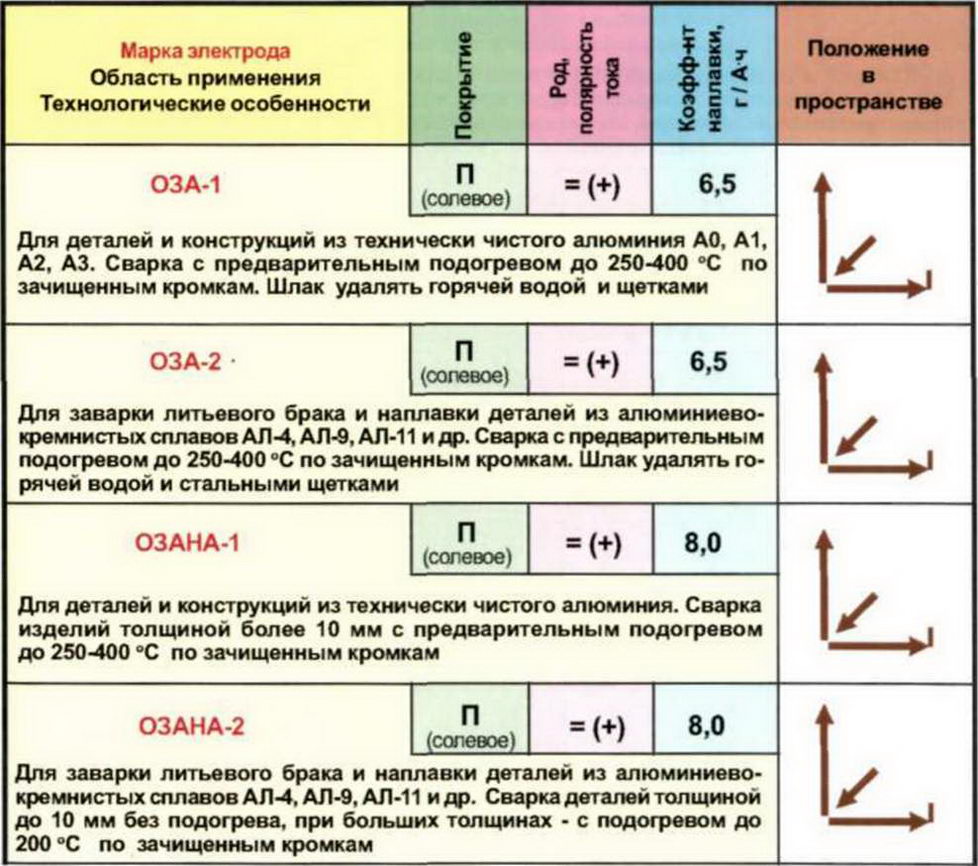 Но на самом деле было всего несколько рандомизированных контролируемых испытаний, и лишь немногие из них были слепыми. Одно исследование много лет назад было простым слепым, что означало, что человек, который оценивал пациента, был слепым, а пациент — нет. Таким образом, пациент знал, что они получают. Есть и другие исследования, в которых все делалось двойным слепым методом — на самом деле мне известно только об одном — но в них не было фиктивного контроля. Под этим я подразумеваю, что вы все были имплантированы, и тогда никто не знал, возбужден он или нет.
Но на самом деле было всего несколько рандомизированных контролируемых испытаний, и лишь немногие из них были слепыми. Одно исследование много лет назад было простым слепым, что означало, что человек, который оценивал пациента, был слепым, а пациент — нет. Таким образом, пациент знал, что они получают. Есть и другие исследования, в которых все делалось двойным слепым методом — на самом деле мне известно только об одном — но в них не было фиктивного контроля. Под этим я подразумеваю, что вы все были имплантированы, и тогда никто не знал, возбужден он или нет.
У нас был фиктивный контроль, в котором мы вставили провод, но не включили его. Но мы пытались заставить пациента думать, что он включен, только из-за эффекта плацебо. Если я убедю вас, что вы что-то получите, многие люди станут лучше только потому, что вы будете убеждены, что вы лучше. Таким образом, это действительно помогает контролировать эффект плацебо. Это действительно единственное испытание, которое было двойным слепым и рандомизированным с фиктивным контролем. Это единственное, что было сделано. Так что на самом деле он предлагает уникальный набор данных, доказательство класса I, и на данный момент единственное в своем роде. Вы действительно не найдете исследование, подобное этому, финансируемое NIH прямо сейчас из-за того, что оно дорогое. Но компании хотят вывести свое устройство на рынок, поэтому мне, как ученому, нравится, что я могу участвовать в этом исследовании, и компания будет платить за это, и мы можем делать некоторые уникальные вещи с исследованиями.
Это единственное, что было сделано. Так что на самом деле он предлагает уникальный набор данных, доказательство класса I, и на данный момент единственное в своем роде. Вы действительно не найдете исследование, подобное этому, финансируемое NIH прямо сейчас из-за того, что оно дорогое. Но компании хотят вывести свое устройство на рынок, поэтому мне, как ученому, нравится, что я могу участвовать в этом исследовании, и компания будет платить за это, и мы можем делать некоторые уникальные вещи с исследованиями.
Был ли какой-то конкретный момент данных или конкретный вывод, который вам запомнился, или, возможно, это было не то, что вы обязательно могли ожидать, отправляясь на испытание?
Нет. Я имею в виду, мы надеялись, что найдем результаты, которые мы видели. Одна из вещей, на которую люди обычно обращают внимание, когда они проводят такого рода исследования, это то, что мы смотрим на количество часов бодрствования, в течение которых пациент проводит время хорошего качества. Мы измеряем это тем, что когда они принимают лекарства, симптомы улучшаются. Мы вызываем это. Когда вы в отключке, это означает, что симптомы действительно проявляются — они ухудшаются. Это называется нерабочее время. Другая часть того, чтобы быть включенным, связана с болезнью Паркинсона, когда вы выходите из положения, в котором у вас очень мало движений, вы медлительны, вы скованны, у вас может быть тремор, дрожь, которая бывает у людей, — это явно не подходит для людей. Когда они включаются, становится лучше, и это уходит в значительной степени. Иногда это не проходит так сильно, как хотелось бы, но может быть заменено чрезмерным движением, называемым дискинезией. Вы переходите от скудости движений, когда вы не в состоянии двигаться, а затем, когда люди становятся более продвинутыми, у них возникают осложнения, так называемая дискинезия.
Мы измеряем это тем, что когда они принимают лекарства, симптомы улучшаются. Мы вызываем это. Когда вы в отключке, это означает, что симптомы действительно проявляются — они ухудшаются. Это называется нерабочее время. Другая часть того, чтобы быть включенным, связана с болезнью Паркинсона, когда вы выходите из положения, в котором у вас очень мало движений, вы медлительны, вы скованны, у вас может быть тремор, дрожь, которая бывает у людей, — это явно не подходит для людей. Когда они включаются, становится лучше, и это уходит в значительной степени. Иногда это не проходит так сильно, как хотелось бы, но может быть заменено чрезмерным движением, называемым дискинезией. Вы переходите от скудости движений, когда вы не в состоянии двигаться, а затем, когда люди становятся более продвинутыми, у них возникают осложнения, так называемая дискинезия.
Что мы искали, так это то, что вовремя? Сколько еще времени они получили без дискинезии? И основной критерий результата заключался в следующем: насколько лучше вы стали после имплантации электрода по сравнению с исходным уровнем по сравнению с 3 месяцами позже.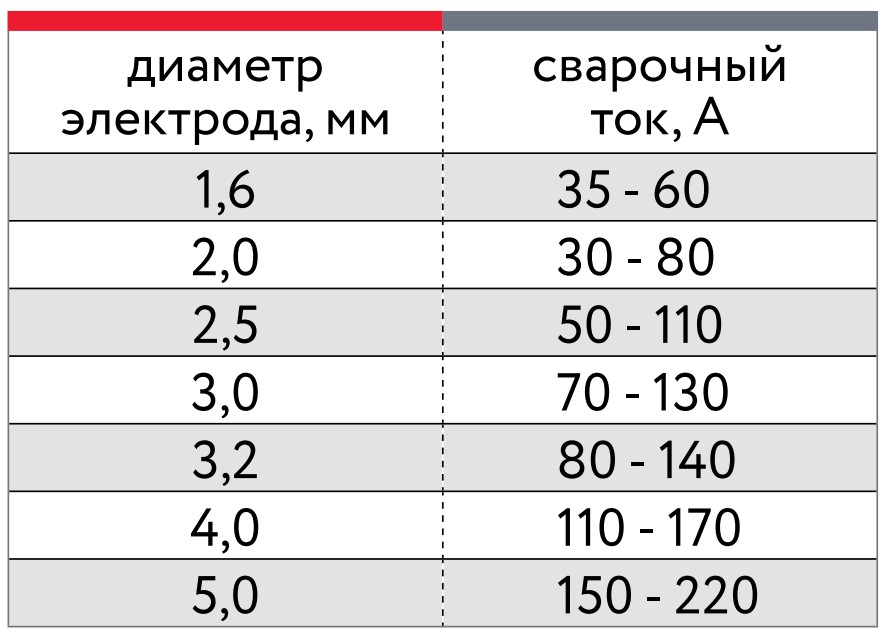 Когда они сделали слепые оценки, мы увидели, что они достигли основного результата, который составил 3 часа. Теперь в большинстве исследований было больше — 4 часа, 4,5 часа. Это исследование сильно отличалось в том смысле, что мы заботились о том, чтобы имплантировать электрод. Сам свинец, просто вставив его, может оказывать влияние на пациентов. И большинство исследований, которые проводились до этого, никогда не рассматривали это. То, на что они смотрели, было во время скрининга, до того, как им имплантировали. Итак, я прошла скрининг, мне вставили имплантат, я выбыла на 3 месяца — как я выглядела до того, как мне вставили имплантат? Если вы посмотрите на этих пациентов с момента скрининга до 3 месяцев после имплантации, то это будет больше похоже на 5 часов. Так что, это немного лучше.
Когда они сделали слепые оценки, мы увидели, что они достигли основного результата, который составил 3 часа. Теперь в большинстве исследований было больше — 4 часа, 4,5 часа. Это исследование сильно отличалось в том смысле, что мы заботились о том, чтобы имплантировать электрод. Сам свинец, просто вставив его, может оказывать влияние на пациентов. И большинство исследований, которые проводились до этого, никогда не рассматривали это. То, на что они смотрели, было во время скрининга, до того, как им имплантировали. Итак, я прошла скрининг, мне вставили имплантат, я выбыла на 3 месяца — как я выглядела до того, как мне вставили имплантат? Если вы посмотрите на этих пациентов с момента скрининга до 3 месяцев после имплантации, то это будет больше похоже на 5 часов. Так что, это немного лучше.
Проблема в том, что все исследования очень разные. Что я оставляю клиницистам, так это то, что когда вы читаете исследование, вам нужно сравнивать яблоки с яблоками, а не яблоки с апельсинами. Мы посмотрели как от скрининга до 3-месячного периода, который длился около 5 часов или более, так и от исходного уровня до 3 месяцев, что на самом деле составило 4,6 часа. Это осложняется тем, что Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) сообщило нам, что если кто-либо в нашей группе увеличил количество принимаемых лекарств после имплантации, даже если они могли улучшить свое лечение вовремя, мы должны были засчитать их своевременность как 0 . Таким образом, несмотря на то, что они стали лучше, правило заключалось в том, что вы должны были назвать это 0. Вот тут-то и появляется число в 3 часа. Суть в том, что лучшее число, вероятно, составляет 4,6 часа, а затем другое число на самом деле показывает скрининг до 3 месяцев, когда они фактически улучшились более чем на 5 часов. Количество времени хорошего качества в течение дня было значительно улучшено.
Мы посмотрели как от скрининга до 3-месячного периода, который длился около 5 часов или более, так и от исходного уровня до 3 месяцев, что на самом деле составило 4,6 часа. Это осложняется тем, что Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) сообщило нам, что если кто-либо в нашей группе увеличил количество принимаемых лекарств после имплантации, даже если они могли улучшить свое лечение вовремя, мы должны были засчитать их своевременность как 0 . Таким образом, несмотря на то, что они стали лучше, правило заключалось в том, что вы должны были назвать это 0. Вот тут-то и появляется число в 3 часа. Суть в том, что лучшее число, вероятно, составляет 4,6 часа, а затем другое число на самом деле показывает скрининг до 3 месяцев, когда они фактически улучшились более чем на 5 часов. Количество времени хорошего качества в течение дня было значительно улучшено.
Если вы посмотрите на цифры скрининга до 3 месяцев, то увидите, что эти цифры не хуже или даже лучше, чем в большинстве исследований на сегодняшний день. Это двойной слепой режим — никто не знает, что происходит. Большинство исследований действительно открытые, поэтому я знаю, что им имплантировали, и они знают, что им имплантировали. Есть небольшой эффект плацебо, своего рода предвзятость, присущая открытым исследованиям. Это было 1 вещь.
Это двойной слепой режим — никто не знает, что происходит. Большинство исследований действительно открытые, поэтому я знаю, что им имплантировали, и они знают, что им имплантировали. Есть небольшой эффект плацебо, своего рода предвзятость, присущая открытым исследованиям. Это было 1 вещь.
Другим важным открытием, на которое обычно обращают внимание люди, является то, как мы оцениваем, когда они не принимают свои лекарства и полностью отказались от лекарств? Насколько им действительно помогает стимуляция? Есть оценочная шкала, которая учитывает ригидность, медлительность, все моторные признаки болезни Паркинсона, и вы записываете их от 0 до 4, и вы получаете число. Затем вы в основном называли все эти цифры и говорили: «Хорошо, мы на 43, когда я не принимаю лекарства, когда я включаю свой стимулятор, до чего я опускаюсь? Какой процент улучшения я вижу?» Это то, что вы ищете. В этом исследовании через 3 месяца в слепой фазе было на 42% лучше. Откровенно говоря, это очень сравнимо с исследованиями, даже открытыми.