Стыковка арматуры в нахлест сколько диаметров снип: Технические особенности нахлеста арматуры при вязке
Содержание
нормы расхода, требования и нюансы
Главная » Строительство »
Верно рассчитанный нахлест арматуры при вязке влияет на итоговое качество конструкции. Надежность такого метода оспорить сложно, однако в процессе работы присутствуют определенные нюансы, при несоблюдении которых результат соединения может оказаться хрупким и недолговечным. Это также может повлиять на скорость затвердевания бетона, что сильно размягчит основание.
Содержание
- Зачем необходимо соблюдать нормы нахлеста арматуры при вязке
- Величина нахлеста при соединении арматуры по СНИП
- Таблица нахлеста арматуры
- Нахлест арматуры при разных условиях
- Нормы расхода арматуры на нахлест
- Важные нюансы и требования для соединения вязкой
Зачем необходимо соблюдать нормы нахлеста арматуры при вязке
При заливке фундамента дома или при возведении любого другого бетонного сооружения (колонны или монолитного блока) насущным остается вопрос прочности и долговечности конструкции. При соблюдении всех строительных норм, дополнительный металлический каркас сильно укрепит конструкцию и сделает ее долговечной, а основание неподверженным влиянию природных условий и времени.
При соблюдении всех строительных норм, дополнительный металлический каркас сильно укрепит конструкцию и сделает ее долговечной, а основание неподверженным влиянию природных условий и времени.
В случае несоблюдения правил, фундамент дома может вскоре обвалиться, что приведет не только к потере большого количества материалов, но и к человеческим жертвам. Это связано с тем, что неверно рассчитанный нахлест арматуры ведет к незатвердеванию бетона в некоторых местах, что приводит к ослабеванию всей конструкции в целом. Для постройки крепкого и надежного каркаса используют несколько способов, в том числе вязку, для которой необходимо использовать нахлест.
Величина нахлеста при соединении арматуры по СНИП
Санитарные Нормы и Правила от 2003 года (сокращенно СНиП) описывают все виды соединений арматур, существующих на данный момент. Стыки внахлест создаются без использования сварочных аппаратов, этим они отличаются от механических (для которых используют муфты и специальное оборудование) и сварных (для которых соответственно нужен сварочный аппарат).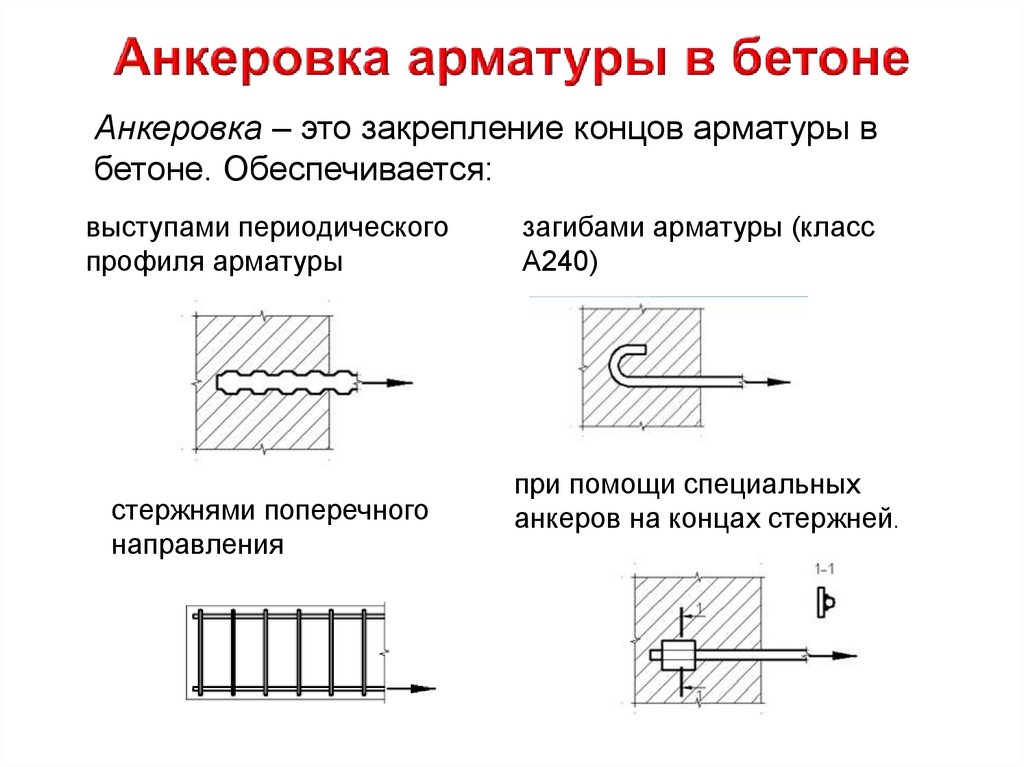 Стыки внахлест существуют трех типов:
Стыки внахлест существуют трех типов:
- Стержни с крюками, лапами (загибами) на концах.
- Стержни, у которых прямой конец (с приваркой или монтажом на пересечении арматур).
- Стержни с прямыми концами (профильные).
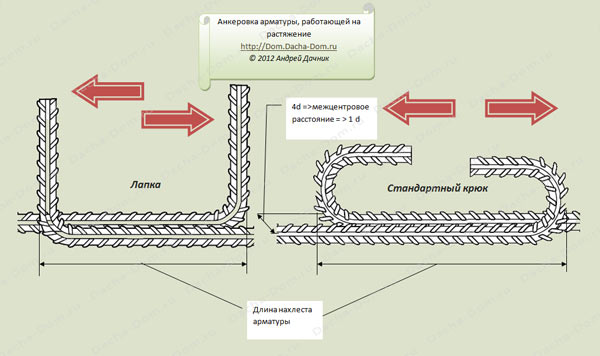
Санитарные Нормы и Правила от 2003 года рекомендуют соединять внахлест арматуры сечением до 40 мм. В свою очередь, мировой аналог строительных норм, а именно ACI 318-05 утверждает максимальное допустимое значение сечения стержней 36 мм. Обусловлено это отсутствием доказательной базы надежности соединений большего диаметра, так как испытания не проводились. Также во время вязки, стоит оставлять определенное свободное пространство вокруг нахлеста.
Надо учитывать, что минимальное расстояние, которое нужно оставить для запаса, как по горизонтали, так и по вертикали составляет 25 мм. Однако, если само сечение арматуры больше 25 мм, то и запас нужно рассчитывать, согласно шагу диаметра. Наибольшим расстоянием между элементами является 8 сечений стержня. Но при использовании в вязке проволоки расстояние сокращается до 4 сечений.
Но при использовании в вязке проволоки расстояние сокращается до 4 сечений.
Не рекомендуется использовать вязку на участках наибольшего давления, так как место соединения не рассчитано на подобные нагрузки, а лишь на крепление арматур и поддержание их в качестве единой конструкции.
Таблица нахлеста арматуры
Величина напуска арматуры в мм
| Диаметр арматурной стали А400 | Величина нахлеста |
| 10мм | 300мм |
| 12мм | 380мм |
| 16мм | 480мм |
| 18мм | 580мм |
| 22мм | 680мм |
| 25мм | 760мм |
| 28мм | 860мм |
| 32мм | 960мм |
| 36мм | 1090мм |
Нахлест арматуры при разных условиях
Места состыковки арматуры и расположение решетки должен определять проектировщик, а не строители. Так как общая картина проекта, а также знание о величине нагрузки в разных местах известны только ему. В противном случае конструкция может быть нарушена.
Так как общая картина проекта, а также знание о величине нагрузки в разных местах известны только ему. В противном случае конструкция может быть нарушена.
Например, во время армирования колонны, следует придерживаться нескольких принципиально важных шагов:
- Выпуск необходимо согнуть на немного большую длину, чем сечение арматуры (для диаметра 16мм — это 20мм).
- Сгибать арматуру необходимо без нагрева, а с помощью специальных средств, которые смогут обеспечить нужный радиус загиба.
- Радиус загиба необходимо указать в проекте и сделать на нем акцент, так как строители вряд ли будут делать это без поручения.
Нормы расхода арматуры на нахлест
Необходимая длина стержней арматуры различается по нескольким критериям:
- Для арматуры работающей на сжатие, необходимая длина будет следующей. Так, для арматур диаметра 6 мм — длина 20-22см; 8мм — длина 20-29см; 10мм — длина 25-36см; 12мм — длина 30-43см; 14мм — длина 35-50см.

- Для арматур работающих на растяжение, требуемая длина нахлеста стержней должна быть больше. Например, для диаметра 6 мм — длина 20-29см; 8мм — длина 27-38см; 10мм — длина 33-48см; 12мм — длина 40-57см; 14мм — длина 46-67см.
Чем выше класс бетона по прочности, тем меньше должна быть длина стержней для нахлеста. Исключениями являются только арматуры 20, 28 и 32 мм. При классе прочности бетона B35 длина стержней должна составлять 655, 920 и 1050 мм соответственно.
Вы соблюдаете нормы нахлеста арматуры при вязке?
Важные нюансы и требования для соединения вязкой
Процесс соединения арматур с помощью проволоки кажется намного более легким, чем вариант со сваркой или же использование спрессованных муфт и специальных аппаратов. Однако он также имеет свои тонкости и нюансы. Надо учитывать, что не стоит соединять арматуры в местах с повышенной нагрузкой (например, углы зданий). Более того, желательно, чтобы в месте вязки нагрузки вообще не было.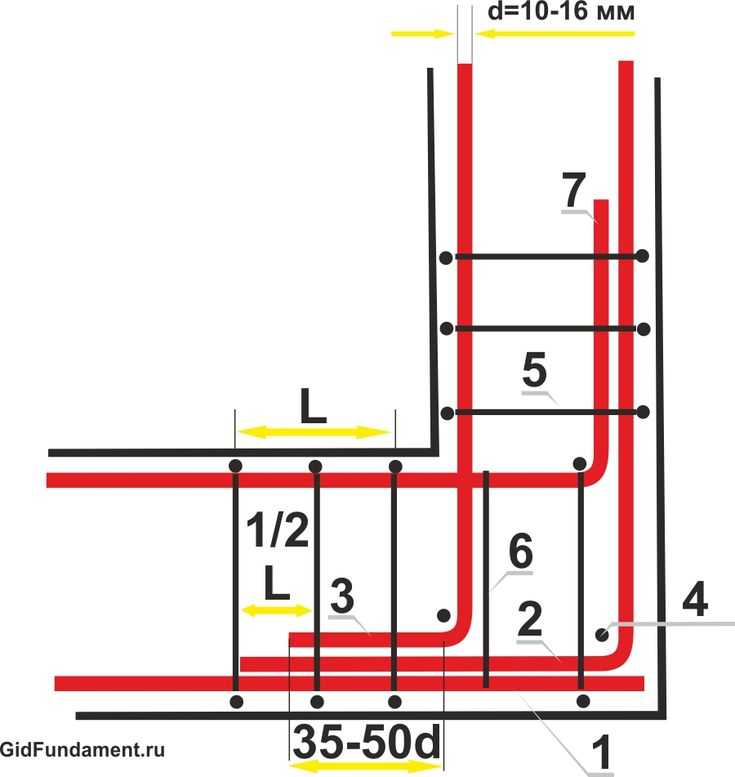 Если же технически нет возможности соблюсти это требование, то стоит пользоваться простой формулой: Размер соединения=90*Сечение используемых прутьев.
Если же технически нет возможности соблюсти это требование, то стоит пользоваться простой формулой: Размер соединения=90*Сечение используемых прутьев.
Также необходимо обращать внимание на основные параметры:
- длину накладки прута;
- местонахождение соединения и особенности данного места;
- расположение нахлестов по отношению друг к другу.
Между соседними местами соединения стрежней арматуры должно быть расстояние, которое можно рассчитать по формуле: Расстояние=1.5*Длину нахлеста, однако получившаяся величина должна быть не меньше 61см.
Также не стоит забывать, что размеры таких соединений регламентированы техническими нормами и нахлест зависит не столько от сечения арматур, сколько от:
- марки бетона, который используется для заливки;
- цели использования соединений;
- класса эксплуатируемой арматуры;
- нагрузки, оказываемой на основание.
Факты, формулы и цифры, изложенные в СНиПе дают представление о том, как именно делать вязку арматур для построения крепкого и надежного каркаса. Эти знания необходимы владельцам дачных участков, которые хотят что-то построить своими силами.
Эти знания необходимы владельцам дачных участков, которые хотят что-то построить своими силами.
Популярное
Нахлест арматуры при армировании: расчет длины и диаметра
Армирование — обязательный этап создания бетонных конструкций. Без каркаса из арматуры просто невозможно создать прочное и долговечное здание или сооружение. При армировании железные прутки соединяют методом сварки или вязки — автоматической или ручной. Важное значение имеет нахлест прутков при соединении арматуры. При недостатке перехлеста прочность конструкции снижается, а делать его с большим запасом экономически невыгодно. Разберемся подробнее — какой делать внахлест при соединении прутков арматуры.
Содержание
- Виды соединения арматуры в каркасе
- Соединение арматуры методом сварки
- Соединение арматуры методом вязки
- Расчет нахлеста по расположению стыка в конструкции
- Расчет нахлеста по марке бетона
- Показатели для сжатой зоны
- Показатели для растянутой зоны
- Важные нюансы и требования для соединения арматуры вязкой
- Нахлест арматуры при разных условиях
- Взаимное расположение перехлестов
- Существенные требования к соединениям арматуры
- Заключение
Виды соединения арматуры в каркасе
При армировании применяют два основных способа соединения узлов. Оба предусмотрены нормативным документом — СНИПом. Он предусматривает следующие правила стыковки прутков при армировании: сварка и вязка. Каждый способ имеет свои особенности, соблюдение которых обязательно.
Оба предусмотрены нормативным документом — СНИПом. Он предусматривает следующие правила стыковки прутков при армировании: сварка и вязка. Каждый способ имеет свои особенности, соблюдение которых обязательно.
Сразу отметим, что соединять сваркой можно не любую арматуру, а только те изделия, которые в маркировке содержат литеру “с”. Причем свариванию подлежат только прутки марки А400 и А500. Состав остальных марок таков, что не позволяет создать прочное соединение с помощью сварочного аппарата.
Соединения арматуры в каркасе
Отметим, что одна из самых популярных марок арматуры А400 может применяться для соединения, но не подходит для наращивания. При сильном нагревании прочность металла этой марки существенно снижается.
Для справки: в местах, где есть нахлест арматуры не рекомендуется применять сварку. И класс стержней не имеет значения. Есть мнение, что при больших нагрузках существенно повышается риск разрыва такого соединения. Отметим, что это мнение распространено в зарубежной технической литературе.
Мнение российских специалистов — если используется дуговая сварка, соединение нахлеста возможно. Но диаметр соединяемых прутков не должен быть больше 25 мм.
Естественно, что особые требования предъявляются и к длине сварного шва. Он напрямую зависит от:
- диаметра прутков;
- класса арматуры.
ГОСТ И СНИП содержат норму:
- для арматуры А400С длина шва кратна восьми диаметрам;
- для А500С — кратна десяти диаметрам.
Пример: если сечение прутка А500С 12 мм, то длина шва сварки не может превышать 120 мм.
Соединение арматуры методом сварки
Сразу отметим, что соединять сваркой можно не любую арматуру, а только те изделия, которые в маркировке содержат литеру “с”. Причем свариванию подлежат только прутки марки А400 и А500. Состав остальных марок таков, что не позволяет создать прочное соединение с помощью сварочного аппарата.
Отметим, что одна из самых популярных марок арматуры А400 может применяться для соединения, но не подходит для наращивания. При сильном нагревании прочность металла этой марки существенно снижается.
Для справки: в местах, где есть нахлест арматуры не рекомендуется применять сварку. И класс стержней не имеет значения. Есть мнение, что при больших нагрузках существенно повышается риск разрыва такого соединения. Отметим, что это мнение распространено в зарубежной технической литературе. Мнение российских специалистов — если используется дуговая сварка, соединение нахлеста возможно. Но диаметр соединяемых прутков не должен быть больше 25 мм.
Естественно, что особые требования предъявляются и к длине сварного шва. Он напрямую зависит от:
- диаметра прутков;
- класса арматуры.
ГОСТ И СНИП содержат норму:
- для арматуры А400С длина шва кратна восьми диаметрам;
- для А500С — кратна десяти диаметрам.

Пример: если сечение прутка А500С 12 мм, то длина шва сварки не может превышать 120 мм.
Соединение арматуры методом вязки
Армирование с помощью вязки — наиболее распространенный способ. Причина в простоте — нет необходимости применять сварочное оборудование, приглашать специалиста. Соединение узлов при армировании проводят специальной вязальной проволокой. Работать с ней умеет многие бетонщики-монолитчики. Да и для каркаса можно применить более дешевые прутки.
Соединение арматуры методом вязки
При связывании прутки прикладывают друг к другу и плотно обвязывают проволокой.
Фиксация арматуры проволокой с нахлестом проводится тремя способами:
- нахлест с фигурными метизами, которые имеют зацепы на концах. Если арматура гладкая, применяют петли и закругленные изделия;
- нахлест с использованием прямых прутков — профиль рифленый;
- нахлест с прямыми прутками — поперечное соединение.

Важно: соединение узлов при армировании без сварки СНИП и ГОСТ допускают только Расчет нахлеста по диаметру арматуры.
Сечение арматуры — величина нормативная. Его подбирают при составлении проекта, согласуясь с требованиями СНИП и ГОСТ. Значение будет иметь плотность монолита и предполагаемые нагрузки. Ориентироваться можно на табличные показатели.
| Диаметр стержней при армировании (для А400) | Кратность диаметров | Длина нахлеста (рекомендуемая величина) (мм) |
|---|---|---|
| 10 | 30 | 300 |
| 12 | 32 | 390 |
| 16 | 30 | 490 |
| 18 | 33 | 590 |
| 22 | 31 | 690 |
| 25 | 30,4 | 770 |
| 28 | 31 | 870 |
| 32 | 30 | 970 |
| 36 | 30,5 | 1090 |
| 40 | 38 | 1590 |
Расчет нахлеста по расположению стыка в конструкции
Плитные и ленточные основания требуют двухконтурного армирования, части которого соединяются вертикальными связями.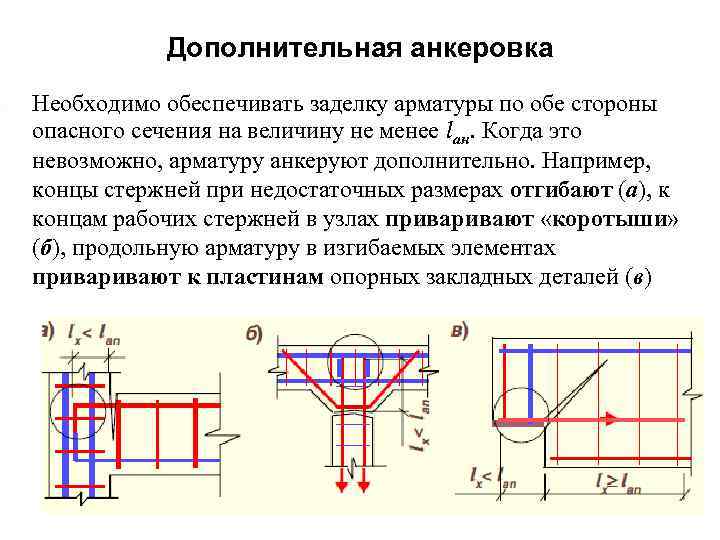 Отметим, что обе части находятся под действием давления — сжатого и растянутого бетона. Отметим, что в обеих зонах требования к длине нахлеста будут различаться.
Отметим, что обе части находятся под действием давления — сжатого и растянутого бетона. Отметим, что в обеих зонах требования к длине нахлеста будут различаться.
Для расчета длины нахлеста оперируют табличными данными.
| Вид бетона/тип фиксации | Горизонтальное (кратно d прутка) | Вертикальное (кратно d прутка) |
|---|---|---|
| Сжатый | 33,8 | 48,3 |
| Растянутый | 47,3 | 67,6 |
Расчет нахлеста по марке бетона
Здесь действует простая зависимость — чем выше показатель марки, тем прочнее конструкция без дополнительного усиления. Это значит, что подойдут прутки меньшего диаметра при армировании, а нахлест при фиксации может быть меньше.
Показатели для сжатой зоны
| Сечение прутка арматуры марки А400/Длина нахлеста по марке бетона | М250 (В20) | М350 (В25) | М400(В30) | М450 (В35) |
|---|---|---|---|---|
| 10 | 360 | 309 | 290 | 260 |
| 12 | 440 | 375 | 365 | 295 |
| 16 | 560 | 490 | 465 | 385 |
| 18 | 650 | 560 | 500 | 440 |
| 22 | 785 | 680 | 580 | 550 |
| 25 | 880 | 780 | 695 | 615 |
| 28 | 995 | 860 | 790 | 680 |
| 32 | 1150 | 975 | 890 | 780 |
| 36 | 1460 | 1230 | 1165 | 990 |
Показатели для растянутой зоны
| Сечение прутка арматуры марки А400(мм)/Длина нахлеста по марке бетона(мм) | М250 (В20) | М350 (В25) | М400(В30) | М450 (В35) |
|---|---|---|---|---|
| 10 | 475 | 410 | 380 | 340 |
| 12 | 590 | 480 | 450 | 395 |
| 16 | 780 | 655 | 595 | 545 |
| 18 | 860 | 740 | 760 | 580 |
| 22 | 1050 | 890 | 890 | 775 |
| 25 | 1190 | 1020 | 950 | 850 |
| 28 | 1330 | 1150 | 1150 | 930 |
| 32 | 1520 | 1300 | 1190 | 1050 |
| 36 | 1890 | 1640 | 1485 | 1315 |
Важные нюансы и требования для соединения арматуры вязкой
Хотя армирование гибкой тонкой арматурой сделать проще, чем сварить ее, важно соблюдать определенные правила. Последние обеспечивают прочность конструктива. Важно учесть:
Последние обеспечивают прочность конструктива. Важно учесть:
- длину нахлеста прутка;
- месторасположение узла армирования и его особенности;
- марку бетона, используемого для создания монолитной конструкции;
- класс арматуры;
- как нахлесты расположены по отношению друг к другу.
Важно: располагать места соединения с нахлестом нельзя на углах здания. Необходимо составить проект армирования таким образом, чтобы расположение узлов было в точках с минимальной нагрузкой.
Требования для соединения арматуры вязкой
Нахлест арматуры при разных условиях
Как уже отмечалось, длина нахлеста зависит от разных факторов и условий. Важные факторы:
- диаметр прутьев. Здесь существует прямая зависимость, чем больше диаметр прутка, тем больше длина нахлеста при армировании. Для примера: если сечение прутка 6 мм, рекомендуемая длина нахлеста составит 250 мм.
 Всегда нужно ориентироваться на 30-40 кратное увеличение сечения для расчета нахлеста при соединении методом вязки;
Всегда нужно ориентироваться на 30-40 кратное увеличение сечения для расчета нахлеста при соединении методом вязки; - минимальное расстояние между узлами — 60 см. Больше можно, а вот меньше не рекомендуется СНИП и ГОСТ.
Нахлест арматуры
Взаимное расположение перехлестов
Прочность бетонной конструкции зависит от правильности армирования. Здесь имеет значение не только длина нахлеста в месте соединения, но и то, как расположены узлы. При составлении проекта их необходимо разнести: подальше друг от друга, применить шахматный порядок. Основные моменты:
- оптимальное расстояние — 140 % от длины нахлеста (допустимо 130-150 %). Это позволяет распределить нагрузку при армировании равномерно, не создавая точек с пограничным давлением;
- одно сечение основания не должно содержать более половины от общего количества перепусков — для периодической арматуры;
- для гладких стержней — не более четверти.
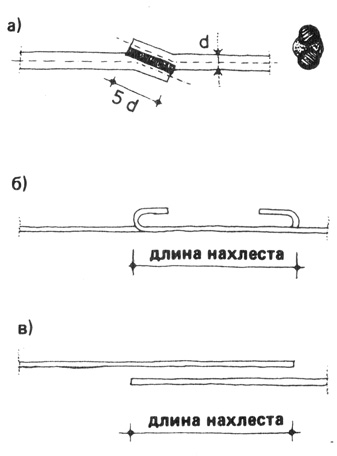
Существенные требования к соединениям арматуры
При связывании узлов армокаркаса важно учесть целый ряд факторов. Отметим, что при возведении обычного ленточного основания можно провести только расчеты по диаметру стержней.
Нюанс: стандартный коэффициент расчета нахлеста при армировании увеличивают до девяносто, если узел предполагается расположить в месте с повышенной нагрузкой, повышенным усилием на изгиб.
Заключение
Армирование — обязательный этап бетонных работ. Чем выше марка применяемого состава, тем ниже требования к усилению. Для создания каркаса используется арматура разного диаметра. Естественно, что прутки необходимо соединять. Основные методы — сварка, вязка узлов с помощью проволоки. Оба способа имеют место на стройке. Первый требует специальных навыков и оборудования. Второй используется чаще.
Оба метода соединения узлов при армировании требуют тщательного расчета длины нахлеста прутков, который позволит обеспечить прочность конструкции. Значение имеет диаметр стержней, метод соединения, марка бетона, расположение узла — в сжатой или растянутой зоне.
Значение имеет диаметр стержней, метод соединения, марка бетона, расположение узла — в сжатой или растянутой зоне.
Соединение арматуры без сварки: способы и типы
Главная
Компания
Соединение арматуры
На сегодняшний день все нормы и правила, а также типы соединений арматуры прописаны в СНиП. Их четкое соблюдение позволяет свести на минимум риски дальнейшей эксплуатации несущих конструкций.
Различают четыре основных вида соединения арматуры: сварочный, соединения «внахлест», обжимные муфты и резьбовые механические соединения.
Виды соединений арматуры
1) Резьбовые механические соединения арматуры Ancon
Инновационное резьбовое соединение арматуры без сварки, благодаря множеству положительных моментов, быстро завоевала доминирующую позицию на соответствующем рынке и повсеместно используется для сооружения многоэтажных зданий, атомных и гидроэлектростанций, мостов и прочих массивных строительных объектов (I и II уровня ответственности).
В отличие от устаревших методов (сварка, вязка), резьбовое соединение продольной арматуры без сварки используется с арматурными стержнями различного диаметра.
Применение резьбовых соединений арматуры в монолитных конструкциях обеспечивает дополнительную прочность, а также экономит металлопрокат (до 20%). Технология также повышает сейсмостойкость и долговечность ЖБИ, одновременно уменьшая нагрузку на фундамент. Такие способы соединения арматуры позволяют сократить время монтажа, заметно снизив общие сроки строительства.
Виды соединения арматуры Ancon
Механическое соединение арматуры Ancon CXL с параллельной резьбой
Муфты соедетельные для арматуры Ancon CXL предназначены для поперечного соединения несущей арматуры. Имеют самые малые габаритные размеры, в тоже время обеспечивают равнопрочное соединение строительной арматуры. Диаметры соединяемой арматуры – 12; 16; 20; 25; 28; 32; 36; 40; 50. для соединения прутков металлопроката разного диаметра возможны переходные муфты для арматуры.
Стыковка арматуры Ancon TT с конической резьбой
Муфты для механического соединения арматуры с конической резьбой разработаны для использования в подавляющем большинстве случаев, в которых необходимо выполнить соединение арматурных стержней. Муфты предназначены для установки на стержни диаметром от 12 до 50мм.
Способ соединения арматуры Ancon MBT
Безрезьбовые механические муфты предназначены для соединения неподготовленной арматуры диаметром от 10 до 40 мм.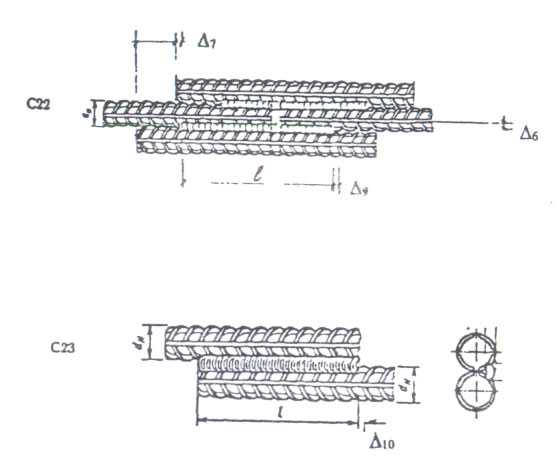 Арматура закрепляется внутри муфты при помощи двух фрикционных накладок и по мере затяжки срезных болтов их конические торцы врезаются в материал стержней. Муфты для стыковки арматуры МВТ особенно удобны в тех случаях, когда арматура уже установлена в конструкции.
Арматура закрепляется внутри муфты при помощи двух фрикционных накладок и по мере затяжки срезных болтов их конические торцы врезаются в материал стержней. Муфты для стыковки арматуры МВТ особенно удобны в тех случаях, когда арматура уже установлена в конструкции.
Муфта для соединения арматуры Ancon RT с накатной цилиндрической резьбой
Одни из самых маленьких муфт в нашем ассортименте, при этом они наиболее рентабельны для применения на крупных проектах. Муфты фирмы Ancon могут упростить проектно-конструкторские и строительные работы и сократить объём арматуры в железобетонных конструкциях.
Технология монтажа
Как правило, установка муфт на арматурные стержни с подготовленной резьбой выполняется на арматурном участке, и стыковые соединения арматуры закрываются пластиковыми колпачками.
Нарезанные концы соединяемых арматурных стержней закрываются пластиковыми или резиновыми защитными колпачками.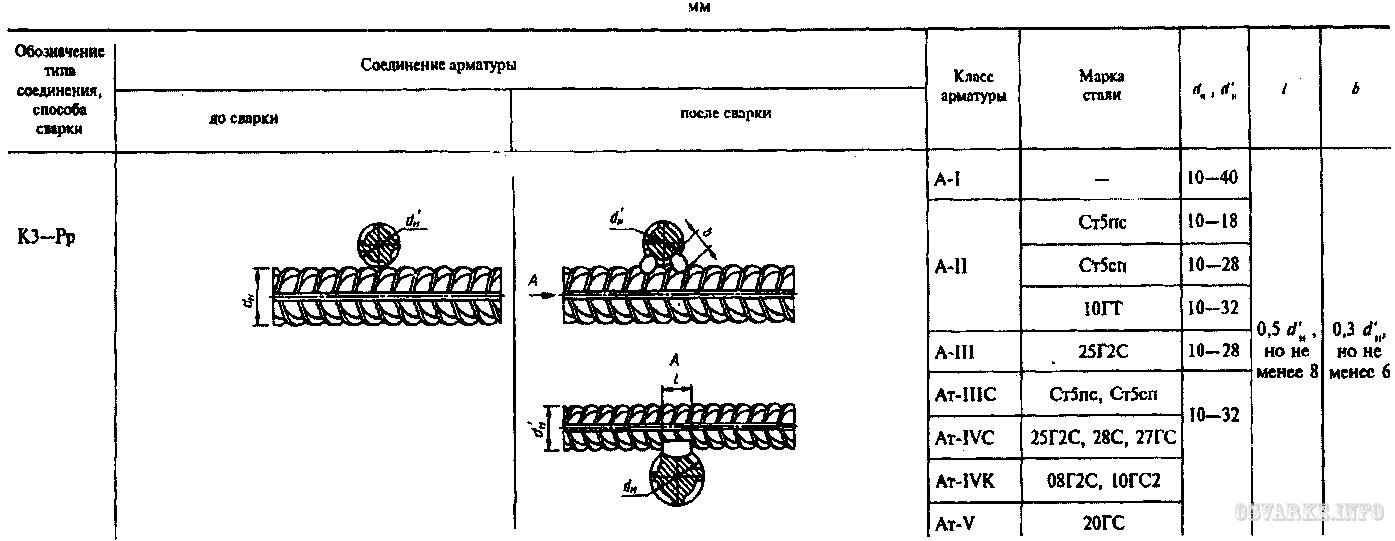
После того, как стержень будет наживлён на муфту, затягивание соединения выполняется ключом с регулированием предельного момента.
2) Соединения «внахлестку»
Соединения «внахлестку» не всегда являются подходящим средством связывания арматуры. Такие способы соединения арматуры (вязка арматурных перепусков) не совсем выгодны – много времени уходит на вязку, что приводит к большему насыщению в бетоне из-за увеличения количества используемых стержней. Соединения «внахлестку» зависимы от бетона при передаче нагрузки. По этой причине любое ухудшение целостности бетона может существенно повлиять на характеристику соединения.
Механические соединения Ancon могут упростить конструкцию и изготовление армированного бетона и уменьшить количество требуемой арматуры. Прочность механического соединения не зависит от бетона, в котором оно размещено, и оно будет сохранять прочность, несмотря на потерю покрытия в результате ударного повреждения или при землетрясении.
3) Обжимные муфты для соединения арматуры
Принцип работы состоит в обжатии муфты из толстостенной стальной трубы и имеют ряд значительных недостатков:
Обжимное муфтовое соединение арматуры нельзя назвать технологией, которая повторяет заявленное качество вне зависимости от условий эксплуатации. Имеются случаи агрессивного поведения гидроприводов, которые норовят пробиться каску работающему персоналу и учинить травму на производстве.
4) Ванная и ванно-шовная сварка
Сущность ванного способа сварки заключается в том, что тепло свариваемым стержням передается не непосредственно под воздействием электрической дуги, а через ванну из жидкого металла. Эта ванна создается за счет расплавления металла электрода и частичного расплавления металла стержней у их торцов. Чтобы предупредить растекание расплавленного металла при сварке, применяют специальные стальные подкладки и накладки, а также инвентарные медные формы. Наплавленный в ванну металл соединяется с расплавленным металлом стержней и образует сварной стыковой шов; при этом стальная подкладка или накладка остается в готовом шве как часть стыка, а медную форму удаляют и используют многократно.
Эта ванна создается за счет расплавления металла электрода и частичного расплавления металла стержней у их торцов. Чтобы предупредить растекание расплавленного металла при сварке, применяют специальные стальные подкладки и накладки, а также инвентарные медные формы. Наплавленный в ванну металл соединяется с расплавленным металлом стержней и образует сварной стыковой шов; при этом стальная подкладка или накладка остается в готовом шве как часть стыка, а медную форму удаляют и используют многократно.
Ванношовная сварка, как способ равнопрочного стыкования строительной арматуры, не имеет перспектив уже в обозримом будущем ввиду наличия непреодолимых качественных ограничений по некоторым присущим ей параметрам, а именно:
- Стоимость соединения;
- Скорость подготовки соединения;
- Объем и способ контроля;
- Квалификация персонала
- Тенденция к использованию термически упрочненной арматуры;
Муфтовые механические соединения Ancon обеспечивают аналогичное качество соединения, не имеют подобных ограничений и позволяют решить строительную задачу любой степени сложности.
Оставить заявку
Сварка арматуры внахлест ГОСТ — О металле
Нормативное основание и типы соединений
Требования снип 52-101-2003 предполагают выполнение условий жесткости для механических и сварных соединений арматурных стержней, а также для соединений прутьев внахлест. Механические соединения арматурных стержней – это резьбовые и прессованные крепления. К строительным операциям, материалам и инструментам применяются не только российские СНИП и ГОСТ – мировая стандартизация ACI 318-05 утверждает нормативное сечение стержня для вязки ≤ 36 мм, в то время как документация внутреннего пользования на российском рынке позволяет увеличить сечение прута до 40 мм. Такое разногласие появилось из-за отсутствия соответствующих задокументированных испытаний арматуры с большим диаметром.
Способы вязания арматурных прутьев
Соединение прутьев арматуры не допускается на локальных участках с превышением допустимых нагрузок и прикладываемых напряжений. Соединение внахлест – это традиционно вязание армостержней мягкой стальной проволокой. Если для армирования фундамента применяется арматура Ø ≤ 25 мм, то практичнее и эффективнее будет использование опрессованных креплений или резьбовых муфт, чтобы повысить безопасность самого соединения и объекта в целом.
Соединение внахлест – это традиционно вязание армостержней мягкой стальной проволокой. Если для армирования фундамента применяется арматура Ø ≤ 25 мм, то практичнее и эффективнее будет использование опрессованных креплений или резьбовых муфт, чтобы повысить безопасность самого соединения и объекта в целом.
К тому же винтовые и опрессованные соединения экономят материал – нахлест прутьев при вязании вызывает перерасход материала ≈ 25%.Строительные нормы и правила № 52-101-2003 регламентируют требования к прочности основания здания – фундамент должен иметь два или более неразрывных контура из арматурных прутьев. Чтобы реализовать это требование на практике, выполняется вязка прутьев внахлест по таким типам:
- Соединение внахлест без сварного шва;
- Соединение сваркой, резьбой или опрессовкой.

Стык внахлест без сварки
Стык без применения сварки чаще всего применяется в индивидуальном строительстве из-за доступности и дешевизны метода. Доступная и недорогая арматура для вязки каркаса – класса A400 AIII. Согласно ACI и СНиП не разрешается стыковать арматуру нахлестом в местах предельных нагрузок и на участках высокой напряженности для арматуры.
Cтыковка арматуры в нахлест
Прочный и долговечный фундамент – это армированный фундамент. Но армирование – операция, требующая точности, и вязание стержней арматуры внахлест или встык требует знания длины прутьев. Лишние сантиметры арматурных прутьев способны деформировать фундамент при прикладываемых боковых нагрузках, нарушить его целостность и общую надежность. И наоборот – правильный монтаж армокаркаса позволит избежать деформирования и растрескивания бетонной ж/б плиты, увеличить срок службы и надежность фундамента. Знание технических особенностей, методов расчета длины прутьев, монтажа стыков и требований снип помогут в строительстве не единожды.
Грамотный нахлест арматуры
Виды соединений
Существует два основных метода крепления арматуры, согласно строительным нормам и правилам (СНиП), а именно пункту 8.3.26 СП 52-101-2003. В нем прописано, что соединение стержней может выполняться следующими типами стыковки:
- Стыковка прутьев арматуры без сварки, внахлест.
- внахлест с использованием деталей с загибами на концах (петли, лапки, крюки), для гладких прутьев используются исключительно петли и крючки;
- внахлест с прямыми концами арматурных прутьев периодического профиля;
- внахлест с прямыми концами арматурных прутьев с фиксацией поперечного типа.
- Механическое и сварное соединение.
- при использовании сварочного аппарата;
- с помощью профессионального механического агрегата.
Требования СНиП указывают на то, что бетонное основание нуждается в установке минимум двух неразрывных каркасов из арматуры. Их делают посредством фиксации стержней внахлест.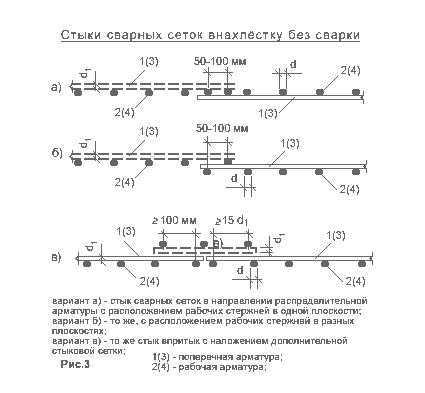 Для частного домостроения подобный способ используется чаще всего. Это связано с тем, что он доступный и дешевый.
Для частного домостроения подобный способ используется чаще всего. Это связано с тем, что он доступный и дешевый.
Обратите внимание! Пункт 8.3.27 гласит, что соединения арматуры внахлест без применения сварки, используется для стержней, рабочее сечение которых не превышает 40 мм. Места с максимальной нагрузкой, не должны фиксироваться внахлест вязкой или сваркой.
Гост 14098-2021 соединения сварные арматуры и закладных изделий железобетонных конструкций. типы, конструкции и размеры (с изменением n 1, с поправкой) от 22 октября 2021 –
ГОСТ 14098-2021
МЕЖГОСУДАРСТВЕННЫЙ СТАНДАРТ
МКС 91.080.40
Дата введения 2021-07-01
Цели, основные принципы и основной порядок работ по межгосударственной стандартизации установлены ГОСТ 1.0-92 “Межгосударственная система стандартизации. Основные положения” и ГОСТ 1.2-2009 “Межгосударственная система стандартизации. Стандарты межгосударственные, правила и рекомендации по межгосударственной стандартизации. Порядок разработки и принятия, применения, обновления и отмены”
Порядок разработки и принятия, применения, обновления и отмены”
Сведения о стандарте
1 РАЗРАБОТАН Научно-исследовательским институтом бетона и железобетона им.А.А.Гвоздева ОАО “НИЦ “Строительство”
2 ВНЕСЕН Техническим комитетом по стандартизации ТК 465 “Строительство”
3 ПРИНЯТ Межгосударственным советом по стандартизации, метрологии и сертификации (протокол от 30 сентября 2021 г. 70-П)
За принятие стандарта проголосовали:
4 Приказом Федерального агентства по техническому регулированию и метрологии от 22 октября 2021 г. N 1374-ст межгосударственный стандарт ГОСТ 14098-2021 введен в действие в качестве национального стандарта Российской Федерации с 01 июля 2021 г.
5 ВЗАМЕН ГОСТ 14098-91
Информация об изменениях к настоящему стандарту публикуется в ежегодном информационном указателе “Национальные стандарты”, а текст изменений и поправок – в ежемесячном информационном указателе “Национальные стандарты”.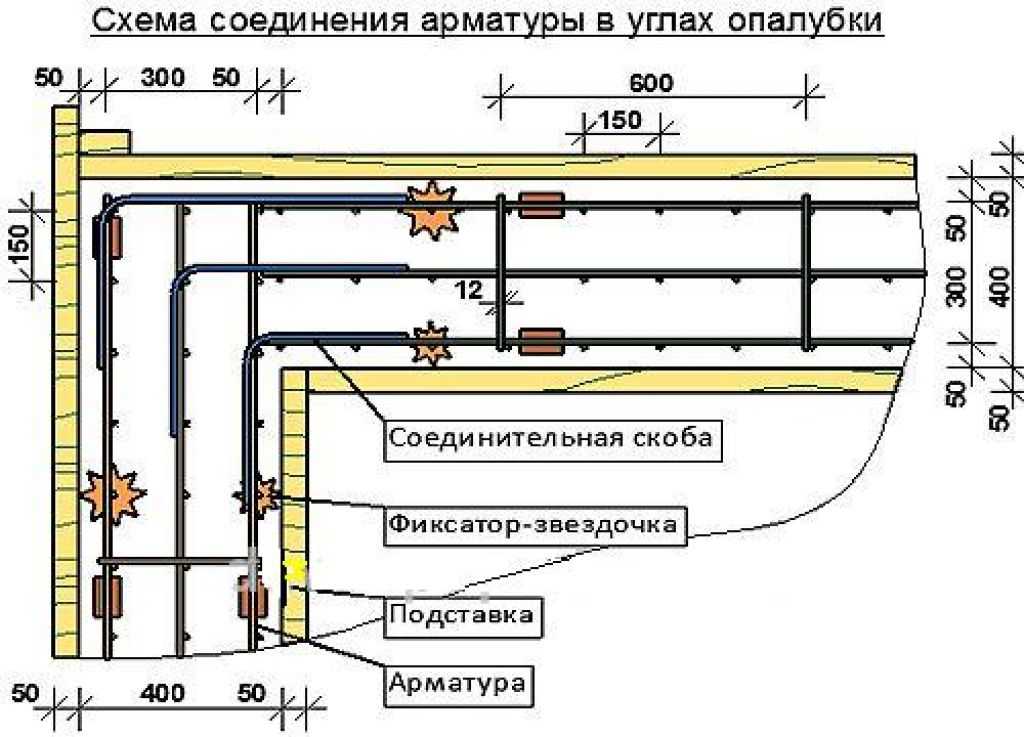 В случае пересмотра (замены) или отмены настоящего стандарта соответствующее уведомление будет опубликовано в ежемесячном информационном указателе “Национальные стандарты”. Соответствующая информация, уведомление и тексты размещаются также в информационной системе общего пользования – на официальном сайте Федерального агентства по техническому регулированию и метрологии в сети Интернет
В случае пересмотра (замены) или отмены настоящего стандарта соответствующее уведомление будет опубликовано в ежемесячном информационном указателе “Национальные стандарты”. Соответствующая информация, уведомление и тексты размещаются также в информационной системе общего пользования – на официальном сайте Федерального агентства по техническому регулированию и метрологии в сети Интернет
ВНЕСЕНО Изменение N 1, утвержденное и введенное в действие приказом Федерального агентства по техническому регулированию и метрологии от 18.04.2021 N 142-ст c 01.09.2021
Изменение N 1 внесено изготовителем базы данных по тексту ИУС N 6, 2021 год
ВНЕСЕНА поправка, опубликованная в ИУС N 9, 2021 год
Поправка внесена изготовителем базы данных
Настоящий стандарт распространяется на сварные соединения стержневой и проволочной арматуры, сварные соединения стержневой арматуры с листовым и фасонным прокатом, выполняемые при изготовлении арматурных и закладных изделий железобетонных конструкций, а также при монтаже сборных и возведении монолитных железобетонных конструкций.
Стандарт устанавливает типы, конструкцию и размеры указанных соединений, выполняемых контактной и дуговой сваркой.
Стандарт не распространяется на сварные соединения закладных изделий, не имеющих анкерных стержней из арматурной стали.
В настоящем стандарте использованы нормативные ссылки на следующие стандарты:
ГОСТ 5264-80 Ручная дуговая сварка. Соединения сварные. Основные типы, конструктивные элементы и размеры
ГОСТ 6727-80 Проволока из низкоуглеродистой стали холоднотянутая для армирования железобетонных конструкций. Технические условия
ГОСТ 8713-79 Сварка под флюсом. Соединения сварные. Основные типы, конструктивные элементы и размеры
ГОСТ 10922-2021* Арматурные и закладные изделия, их сварные, вязаные и механические соединения для железобетонных конструкций. Общие технические условия
________________
* На территории Российской Федерации действует ГОСТ Р 57997-2021 “Арматурные и закладные изделия сварные, соединения сварные арматуры и закладных изделий железобетонных конструкций.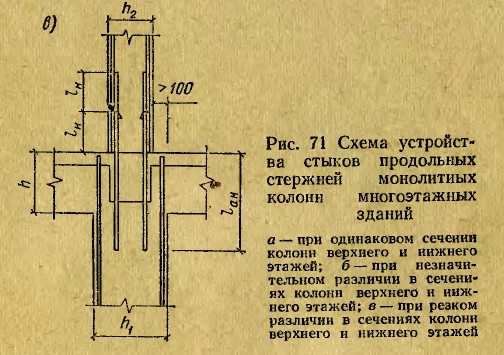 Общие технические условия”.
Общие технические условия”.
ГОСТ 14771-76 Дуговая сварка в защитном газе. Соединения сварные. Основные типы, конструктивные элементы и размеры
ГОСТ 27772-88 Прокат для строительных конструкций. Общие технические требования
ГОСТ 34028-2021 Прокат арматурный для железобетонных конструкций. Технические условия
Примечание – При пользовании настоящим стандартом целесообразно проверить действие ссылочных стандартов в информационной системе общего пользования – на официальном сайте Федерального агентства по техническому регулированию и метрологии в сети Интернет или по ежегодному информационному указателю “Национальные стандарты”, который опубликован по состоянию на 1 января текущего года, и по выпускам ежемесячного информационного указателя “Национальные стандарты” за текущий год. Если ссылочный стандарт заменен (изменен), то при пользовании настоящим стандартом следует руководствоваться заменяющим (измененным) стандартом. Если ссылочный стандарт отменен без замены, то положение, в котором дана ссылка на него, применяется в части, не затрагивающей эту ссылку.
Если ссылочный стандарт отменен без замены, то положение, в котором дана ссылка на него, применяется в части, не затрагивающей эту ссылку.
(Измененная редакция, Изм. N 1), (Поправка. ИУС N 9-2021).
В настоящем стандарте использованы термины по ГОСТ 10922.
(Измененная редакция, Изм. N 1).
4.1 Обозначения типов сварных соединений и способов их сварки приведены в таблице 1.
Таблица 1 – Обозначения типов сварных соединений и способов их сварки
4.2 Условное обозначение сварного соединения имеет следующую структуру
Пример условного обозначения стыкового соединения, выполненного ванно-шовной сваркой на стальной скобе-накладке, положение стержней горизонтальное:
Как расположить друг относительно друга арматурные перепуски
Для увеличения прочности силового каркаса фундамента очень важно правильно располагать нахлесты арматуры относительно друг друга в обеих плоскостях тела бетона.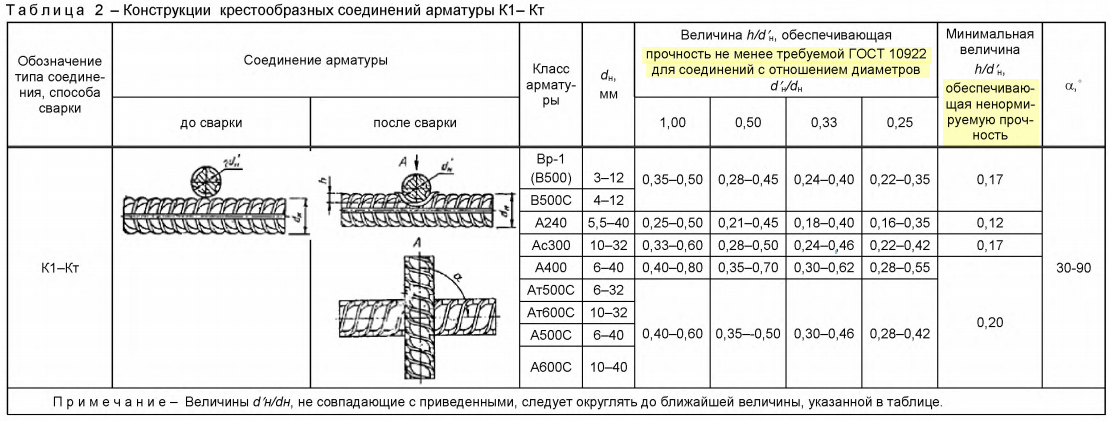 СНиП и ACI рекомендуют разносить соединения, таким образом, чтоб в одном сечении было не более 50% перепусков.
СНиП и ACI рекомендуют разносить соединения, таким образом, чтоб в одном сечении было не более 50% перепусков.
Взаимное расположение арматурных перепусков в теле бетона
Если центры нахлеста вязаной арматуры находятся в пределах указанной величины, то считается, что соединения стержней располагается в одном сечении.
Согласно нормам ACI 318-05 взаимное расположение стыковочных соединений должно находиться на расстоянии не менее 61 сантиметра. Если дистанция будет не соблюдена, то повышается вероятность деформации бетонного монолитного основания от нагрузок, оказываемых на него в процессе возведения здания и его последующей эксплуатации.
Какую делать величину нахлеста арматуры при вязке
Поскольку вязка арматуры внахлест определяется технической документацией, то там четко указана протяженность стыковочных соединений. При этом величины могут колебаться не только от диаметра используемых прутов, но и от таких показателей как:
- Характер нагрузки;
- Марка бетона;
- Класс арматурной стали;
- Мест соединения;
- Назначения ЖБИ (горизонтальные плиты, балки или вертикальные колонны, пилоны и монолитные стены).

Сращивание арматурных стержней при выполнении нахлеста
В целом же протяженность нахлеста прутов арматуры при вязке определяется влиянием усилий, возникающих в стержнях, воспринимаемых сил сцеплением с бетоном, воздействующими по всей длине стыка, и силами, оказывающими сопротивления в анкеровке армирующих прутов.
Основополагающим критерием при определении длинны напуска арматуры при вязке, берется ее диаметр.
Для удобства расчетов нахлеста армирующих стержней при вязке силового каркаса монолитного фундамента предлагаем воспользоваться таблицей с указанными величинами диаметра и их напуска. Практически все величины сводятся к 30-ти кратному диаметру применяемых стержней.
| Величина напуска арматуры в диаметрах | ||
|---|---|---|
| Диаметр арматурной стали А400, мм | Величина нахлеста | |
| в диаметрах | в мм | |
| 10 | 30 | 300 мм |
| 12 | 31,6 | 380 мм |
| 16 | 30 | 480 мм |
| 18 | 32,2 | 580 мм |
| 22 | 30,9 | 680 мм |
| 25 | 30,4 | 760 мм |
| 28 | 30,7 | 860 мм |
| 32 | 30 | 960 мм |
| 36 | 30,3 | 1090 мм |
В зависимости от нагрузок и назначения железобетонных изделий длина нахлестных соединений стержневой стали изменяется в сторону увеличения:
| Напуск арматуры в зависимости от назначения ЖБИ | ||
|---|---|---|
| Вид нагрузки | Назначение ЖБИ | |
| Горизонтальное использование, в диаметрах | Вертикальное использование, в диаметрах | |
| В сжатом бетоне | 33,8 ᴓ | 48,3 ᴓ |
| В растянутом бетоне | 47,3 ᴓ | 67,6 ᴓ |
В зависимости от марки бетона и характера нагрузки, применяемого для заливки монолитной ленты фундамента и прочих железобетонных элементов, минимальные рекомендуемые величины перепуска арматуры в процессе вязки будут следующими:
| Для сжатого бетона | ||||
|---|---|---|---|---|
| Диаметр армирующей стали А400 используемой в сжатом бетоне, мм | Длина нахлеста армирующих стержней для марок бетона (класс прочности бетона), в мм | |||
| М250 (В20) | М350 (В25) | М400 (В30) | М450 (В35) | |
| 10 | 355 | 305 | 280 | 250 |
| 12 | 430 | 365 | 335 | 295 |
| 16 | 570 | 490 | 445 | 395 |
| 18 | 640 | 550 | 500 | 445 |
| 22 | 785 | 670 | 560 | 545 |
| 25 | 890 | 765 | 695 | 615 |
| 28 | 995 | 855 | 780 | 690 |
| 32 | 1140 | 975 | 890 | 790 |
| 36 | 1420 | 1220 | 1155 | 985 |
| Для растянутого бетона | ||||
|---|---|---|---|---|
| Диаметр армирующей стали А400 используемой в растянутом бетоне, мм | Длина нахлеста армирующих стержней для марок бетона (класс прочности бетона), в мм | |||
| М250 (В20) | М350 (В25) | М400 (В30) | М450 (В35) | |
| 10 | 475 | 410 | 370 | 330 |
| 12 | 570 | 490 | 445 | 395 |
| 16 | 760 | 650 | 595 | 525 |
| 18 | 855 | 730 | 745 | 590 |
| 22 | 1045 | 895 | 895 | 275 |
| 25 | 1185 | 1015 | 930 | 820 |
| 28 | 1325 | 1140 | 1040 | 920 |
| 32 | 1515 | 1300 | 1185 | 1050 |
| 36 | 1895 | 1625 | 1485 | 1315 |
Контактный метод
Самым распространенным методом сваривания листовых материалов является соединение их внахлест.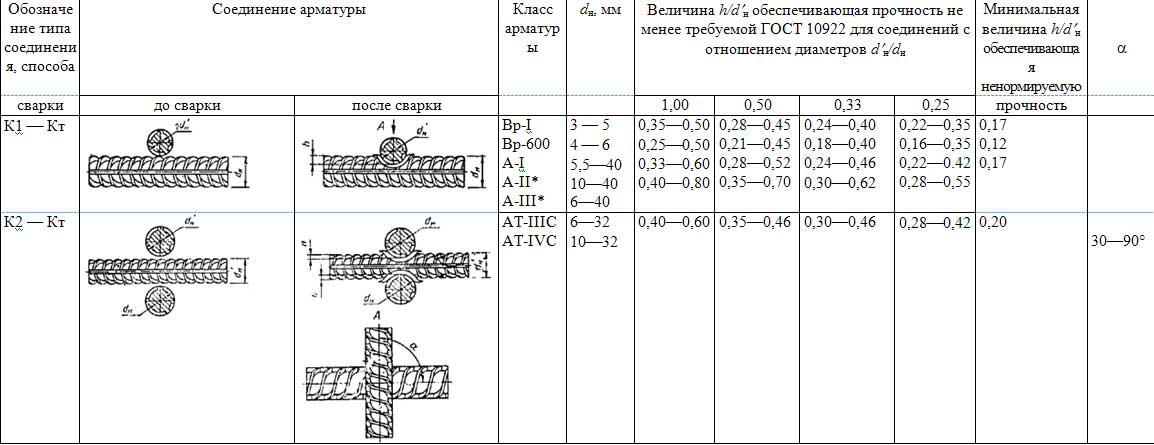 Его осуществляют с помощью рельефов (специальных выступов). Обычно применяют рельефы сферической формы. Рельефная сварка относится к разновидностям контактного метода.
Его осуществляют с помощью рельефов (специальных выступов). Обычно применяют рельефы сферической формы. Рельефная сварка относится к разновидностям контактного метода.
При сварочном процессе внахлест рельефы формуют с применением холодной штамповки, что вызывает образование лунки. Если использовать материалы с высокой пластичностью, то можно получить рельефы любой сложности. Если рельефы получить затруднительно по каким-либо причинам, то можно использовать специальные вставки.
По сравнению с контактным сварочным процессом рельефный метод имеет некоторые отличия. Так, сварное соединение получается не за счет плавления металла, а за счет пластической деформации.
Контактная сварка в этом плане более требовательная, в ней сварочные точки не могут располагаться слишком близко к краю стыка. Между собой они тоже на должны находиться близко из-за шунтирующих токов.
Несмотря на это, контактная сварка внахлест очень распространена в автомобилестроении и приборостроении, широко применяется в изготовлении бытовой техники.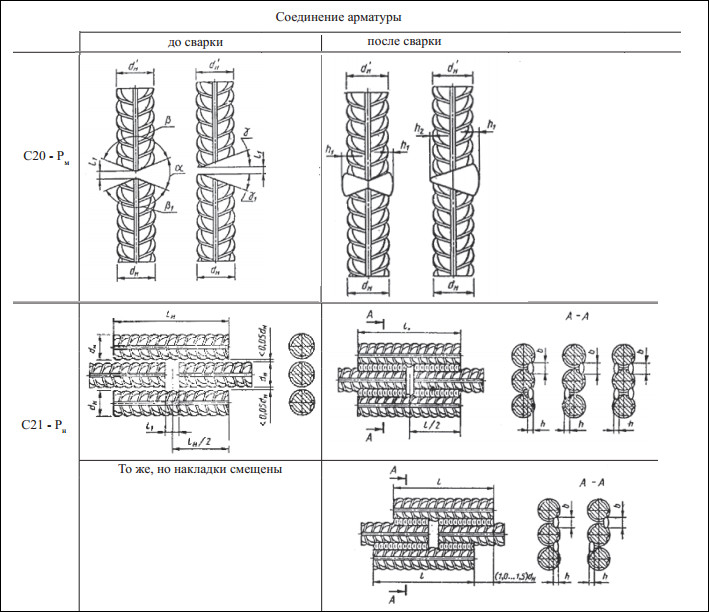 Сам принцип действия контактной сварки предполагает нахлесточное соединение.
Сам принцип действия контактной сварки предполагает нахлесточное соединение.
Местонахождение соединений арматуры внахлест
Нормативные документы не разрешают располагать участки соединения арматуры ввязкой в местах предельных нагрузок и напряжений. Все стыки стержней рекомендуется располагать в железобетонных конструкциях с ненагруженными участками и без приложения напряжений. Для ленточных монолитных фундаментов участки перепуска концов прутьев нужно размещать в локальных участках с без приложения крутящих и изгибающих сил, или с минимальным их вектором. При невозможности выполнения этих требований, длина перепуска армостержней принимается как 90 Ø соединяемой арматуры.
Расположение арматуры при вязке
Общая длина всех вязаных перепусков в каркасе зависит от приложенных усилий к прутьям, уровня сцепления с бетоном и напряжений, возникающих по протяженности соединения, а также сил сопротивления в перехлестах армопрутьев. Главный параметр при расчете длины перепуска соединяемой арматуры – диаметр стержня.
Калькулятор
Таблица ниже позволяет без сложных расчетов определить нахлест армирующих прутьев при монтаже армирующего фундаментного каркаса. Почти все значения в таблице приводятся к Ø 30 связываемых армирующих стержней.
| Перепуск стержней в Ø | ||
| Ø стали класса А 400, мм | Перепуск | |
| в Ø | в мм | |
| 10 | 30 | 300 |
| 12 | 31,6 | 380 |
| 16 | 30 | 480 |
| 18 | 32,2 | 580 |
| 22 | 30,9 | 680 |
| 25 | 30,4 | 760 |
| 28 | 30,7 | 860 |
| 32 | 30 | 960 |
| 36 | 30,3 | 1090 |
Чтобы повысить прочность армокаркаса основания дома, нахлесты в арматуре необходимо правильно располагать по отношению друг к другу.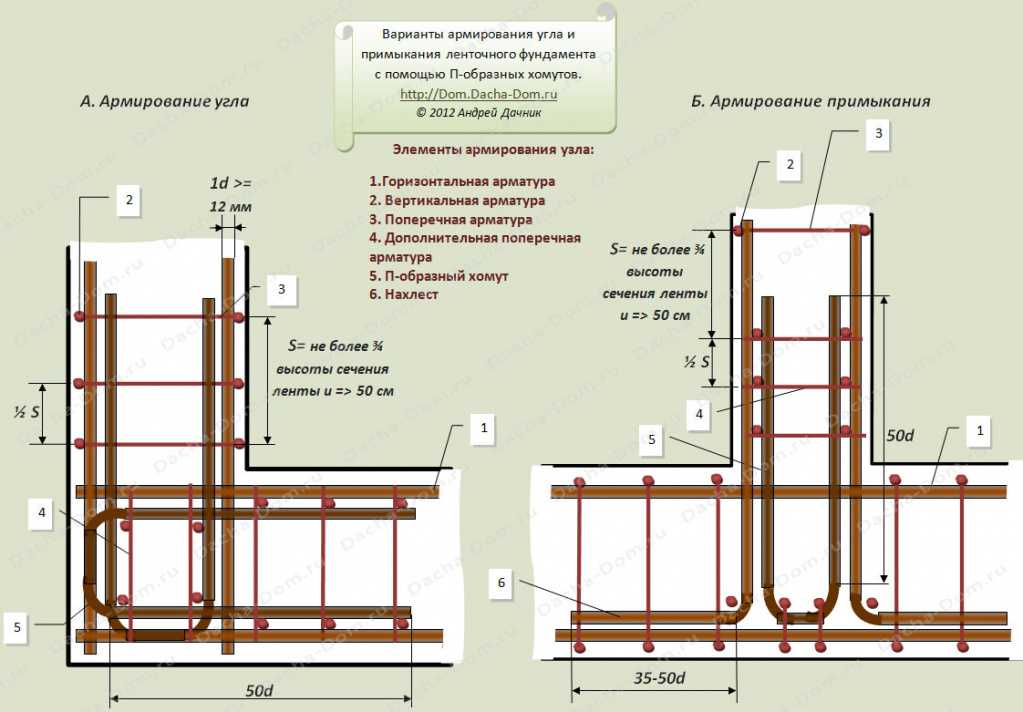 причем контролировать размещение и в горизонтальной, и в вертикальной плоскости в бетоне. Российские и международные нормы и правила рекомендуют по этому поводу делать разнос связок, чтобы в одном разрезе находилось не более 50% нахлестов. Расстояние разнесения, определенное СНиП и ACI, не должно быть больше 130% всей длины стыков армирующих прутьев.
причем контролировать размещение и в горизонтальной, и в вертикальной плоскости в бетоне. Российские и международные нормы и правила рекомендуют по этому поводу делать разнос связок, чтобы в одном разрезе находилось не более 50% нахлестов. Расстояние разнесения, определенное СНиП и ACI, не должно быть больше 130% всей длины стыков армирующих прутьев.
Как располагать нахлесты прутьев
Международные требования ACI 318-05 определяют разнесение стыков на расстояние ≥ 61 см. При превышении этого значения вероятность деформирования бетонного фундамента от напряжений и нагрузок значительно возрастает.
источник
Нахлест при разных условиях
Так какой же нахлест арматуры при вязке? Какие есть точные данные? Начнем с рассмотрения примеров. Первый фактор, от которого зависит нахлест – это диаметр прутьев. Наблюдается следующая закономерность: чем больше диаметр используемой арматуры, тем больше становится нахлест.
Итак, чтобы упростить задачу, используем специальную таблицу, где указан, какой нахлест используется для прутьев разного диаметра.
| Диаметр используемой арматуры А400 (мм) | Количество диаметров | Предполагаемый нахлест (мм) |
| 10 | 30 | 300 |
| 12 | 31,6 | 380 |
| 16 | 30 | 480 |
| 18 | 32,2 | 580 |
| 22 | 30,9 | 680 |
| 25 | 30,4 | 760 |
| 28 | 30,7 | 860 |
| 32 | 30 | 960 |
| 36 | 30,3 | 1090 |
| 40 | 38 | 1580 |
С этими данными каждый сможет выполнить работу правильно. Но есть еще одна таблица, указывающая на нахлест при использовании сжатого бетона. Он зависит от класса используемого бетона. При этом чем выше класс, тем разбежка стыков арматуры меньше.
| Сечение арматуры А400, которая используется для работы (мм) | Длина нахлеста, в зависимости от марки бетона (мм) | |||
| В20 (М250) | В25 (М350) | В30 (М400) | В35 (М450) | |
| 10 | 355 | 305 | 280 | 250 |
| 12 | 430 | 365 | 355 | 295 |
| 16 | 570 | 490 | 455 | 395 |
| 18 | 640 | 550 | 500 | 445 |
| 22 | 785 | 670 | 560 | 545 |
| 25 | 890 | 765 | 695 | 615 |
| 28 | 995 | 855 | 780 | 690 |
| 32 | 1140 | 975 | 890 | 790 |
| 36 | 1420 | 1220 | 1155 | 985 |
Что касается растянутой зоны бетона, то в отличие от сжатой зоны, нахлест будет еще больше.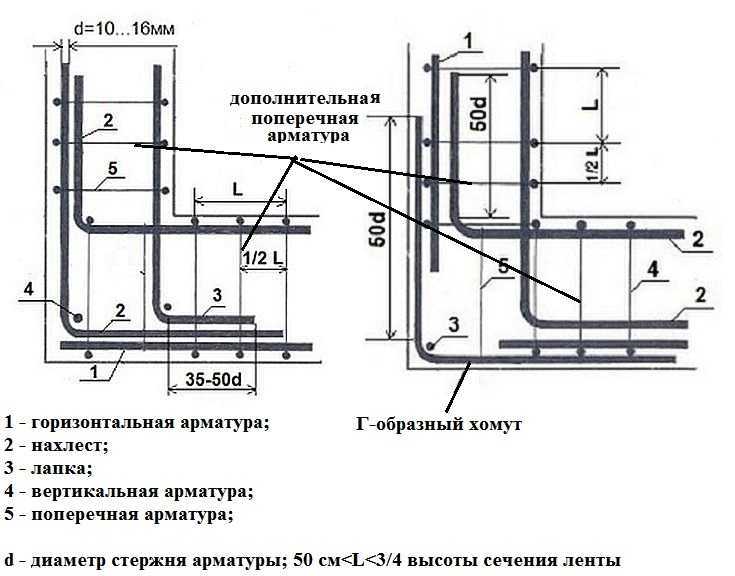 Как и в предыдущем случае, с увеличением марки раствора длина уменьшается.
Как и в предыдущем случае, с увеличением марки раствора длина уменьшается.
| Сечение арматуры А400, которая используется для работы (мм) | Длина нахлеста, в зависимости от марки бетона (мм) | |||
| В20 (М250) | В25 (М350) | В30 (М400) | В35 (М450) | |
| 10 | 475 | 410 | 370 | 330 |
| 12 | 570 | 490 | 445 | 395 |
| 16 | 760 | 650 | 595 | 525 |
| 18 | 855 | 730 | 745 | 590 |
| 22 | 1045 | 895 | 895 | 775 |
| 25 | 1185 | 1015 | 930 | 820 |
| 28 | 1325 | 1140 | 1140 | 920 |
| 32 | 1515 | 1300 | 1185 | 1050 |
| 36 | 1895 | 1625 | 1485 | 1315 |
Если правильно расположить нахлест друг относительно друга и сделать его нужной длины, то скелет основания получит значительные увеличения прочности.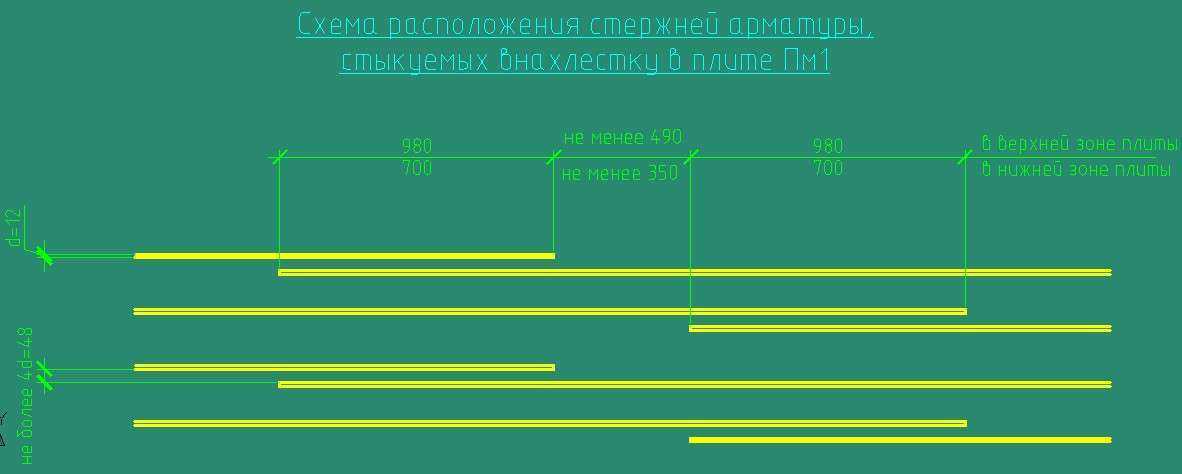 Соединения равномерно распределяются по всей конструкции.
Соединения равномерно распределяются по всей конструкции.
Согласно нормам и правилам (СНиП), минимальное расстояние между соединением должно составлять 61 см. Больше – лучше. Если не соблюдать эту дистанцию, то риск, что конструкция при сильных нагрузках и в ходе эксплуатации будет деформироваться, возрастает. Остается следовать рекомендациям, для создания качественного армирования.
источник
Во время армирования фундамента или изготовления любого из видов армопояса практически у каждого человека возникает вопрос о том, какой должна быть длина нахлеста, и каким образом правильно его выполнить. Действительно, это имеет большое значение. Верно выполненная стыковка стальных прутьев делает более прочным соединение арматуры.
Нахлест арматуры при вязке – это самый простой и при этом по-настоящему надежный вариант соединения арматуры
Сварка арматуры согласно гост
Арматурный каркас– это система соединенных между собой элементов. В железобетонных изделиях металл воспринимает нагрузки на сжатие и растяжение.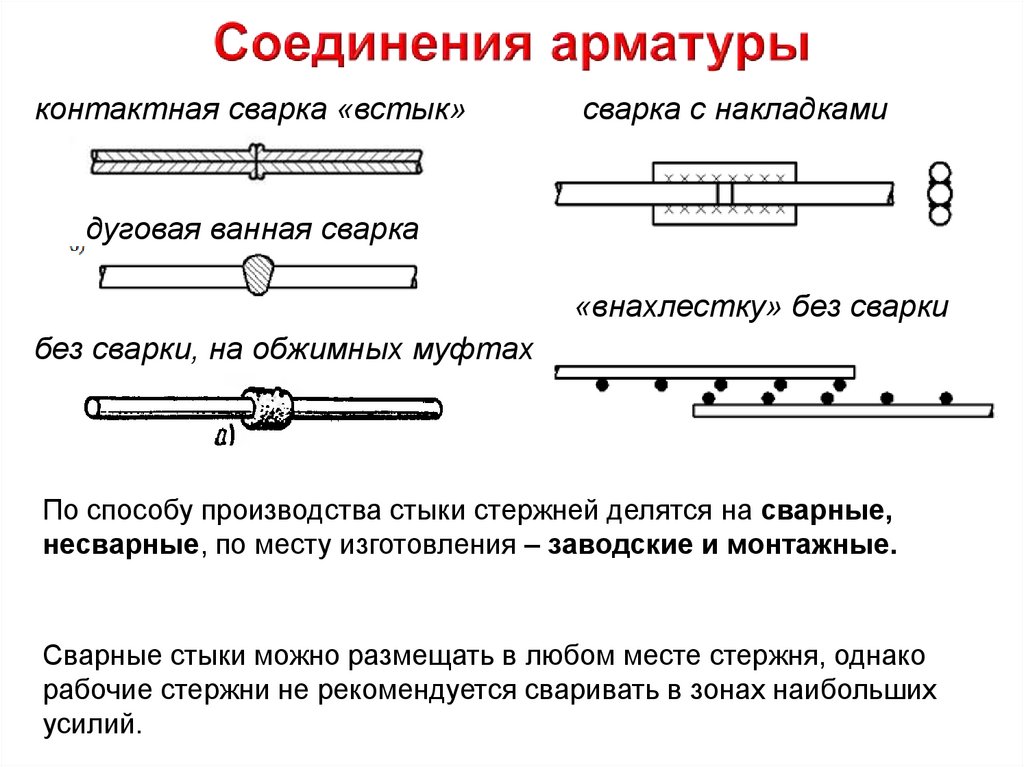 Сварка арматуры (ГОСТ 14098-91) позволяет создать каркас, который жестче вязаного. Проволочное сцепление реализуется в особых случаях.
Сварка арматуры (ГОСТ 14098-91) позволяет создать каркас, который жестче вязаного. Проволочное сцепление реализуется в особых случаях.
Сварка арматуры контактной сваркой, впрочем, как и контактно- точечная сварка арматуры проводится в производственных условиях на специальном профессиональном оборудовании.
Домашний мастер при реализации работ использует более простые устройства на ручном или автоматическом управлении.
Контактная сварка арматуры необходима при строительстве крупных объектов городской и промышленной застройки.
Правильное армирование увеличивает прочность возводимой конструкции на 245%, на фоне повышения стойкости к механическому воздействию – на 150%
Соединение внахлест вязанием
Дешевый и распространенный класс арматуры для соединений без сварки – А400 АIII. Стыки скрепляются вязальной проволокой, к местам вязки предъявляются особые требования.
Стыки скрепляются вязальной проволокой, к местам вязки предъявляются особые требования.
Анкеровка или нахлест арматуры при вязке таблица значений которого приведена ниже для вязки в бетоне марки BIO с прочностью 560 кг/см 2 , предполагает использование определенных марок и классов армостержней с определенным типом металлообработки для определенных диаметров:
Работа арматуры при сжатии и растяжении
Механическая стыковка прутьев в каркасе для ж/б изделий проводится один из следующих способов:
- Наложением прямых стержней друг на друга;
- Нахлест прута с прямым концом со сваркой или механическим креплением на всем перепуске поперечных стержней;
- Механическое и сварное крепление стержней с загнутыми в виде крючков, петель и лап законцовками.
Применение гладкой арматуры требует вязать ее внахлест или сваривать с поперечными прутьями каркаса.
Требования к вязке прутьев внахлест:
- Необходимо вязать стержни с соблюдением длины наложения прутьев;
- Соблюдать нахождение мест вязки в бетоне и перепусков арматуры по отношению друг к другу;
Соблюдение требований СНиП позволит эксплуатировать прочные ж/ плиты в фундаментах с большим и гарантированным сроком службы.
Способы ручной вязки арматуры
Соединение нахлеста арматурных стержней сваркой
Для дачного строительства сварка нахлеста арматуры считается дорогим удовольствием, по причине высокой стоимости металлических стержней марки А400С или А500С. Они относятся к свариваемому классу. Что существенно повышает стоимость материалов. Использовать пруты без индекса «С», например: распространенный класс A400 AIII, недопустимо, так как при нагревании металл значительно теряет свою прочность и коррозионную стойкость.
Тем не менее, если Вы решили использовать стержни свариваемого класса (А400С, А500С, В500С), их соединения следует сваривать электродами 4…5 миллиметрового диаметра. Протяженность сварочного шва и самого нахлеста зависит от используемого класса арматуры.
| Протяженность сварочного шва при нахлесте | |
|---|---|
| Класс арматурных стержней | Протяженность сварного шва нахлеста в диаметрах соединяемой арматуры |
| А400С | 8 ᴓ |
| А500С | 10 ᴓ |
| В500С | 10 ᴓ |
Исходя из приведенных данных видно, что при использовании при вязке стальных прутов класса В400С величина нахлеста, соответственно и сварного шва, составит 10 диаметров свариваемой арматуры. Если для силового каркаса фундамента взяты стержни ᴓ12 мм, то протяженность шва составит 120 мм, что, по сути, будет соответствовать ГОСТу 14098 и 10922.
Согласно американским нормам нельзя сваривать перекрестия арматурных стержней. Действующие нагрузки на основание могут вызвать возможные разрывы, как самих прутьев, так и мест их соединения.
Соединение прутьев методом сварки
Нахлест стержней методом сварки используется исключительно с арматурой марки А400С и А500С.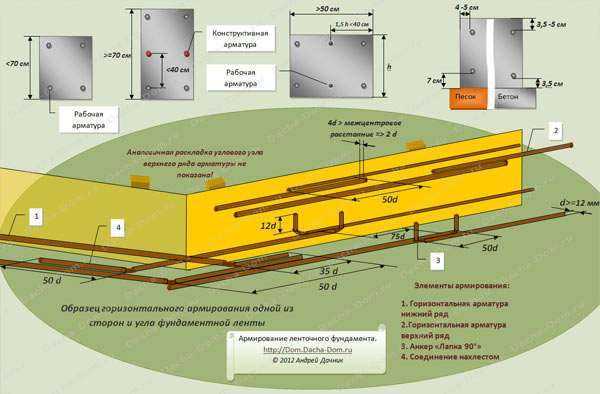 Только эти марки считаются свариваемыми. Это сказывается и на стоимости изделий, которая выше обычных. Одним из распространенных классов является класс А400. Но сращивание изделий ими недопустимо. Нагреваясь, материал становится менее прочным и теряет свою устойчивость к коррозии.
Только эти марки считаются свариваемыми. Это сказывается и на стоимости изделий, которая выше обычных. Одним из распространенных классов является класс А400. Но сращивание изделий ими недопустимо. Нагреваясь, материал становится менее прочным и теряет свою устойчивость к коррозии.
В местах, где есть перехлест арматуры, сваривание запрещается, несмотря на класс стержней. Почему? Если верить зарубежным источникам, то есть большая вероятность разрыва места соединения, если на него будут воздействовать большие нагрузки. Что касается российских правил, то мнение следующее: использовать дуговую электросварку для стыковки разрешается, если размер диаметров не будет превышать 25 мм.
Важно! Длина сварочного шва напрямую зависит от класса арматурного прута и его диаметра. Для работы используют электроды, сечение которых от 4 до 5 мм. Требования, регламентированные в ГОСТах 14098 и 10922, сообщают, что делать нахлест методом сварки можно длиной меньше 10 диаметров арматурных прутьев, используемых для работ.
Соединение стержней сваркой
Нахлест арматуры с использованием сварки допускается только со стержнями марок А400С и А500С. Арматура этого класса считается свариваемой. Но стоимость таких стержней достаточно высока. Самый же распространенный класс — А400. Но его использование недопустимо, так как при его нагревании заметно сокращается прочность и устойчивость к коррозии.
Запрещается сваривать места, где есть перехлест арматуры, независимо от класса последней. Существует вероятность разрывов стержней при воздействии на них больших нагрузок. Так говорят зарубежные источники. В российских правилах разрешается использование дуговой электросварки этих мест, но размер диаметров не должен превышать 2,5 см.
Арматуру запрещено соединять в местах максимального напряжения стержней и зонах приложения (концентрированного) нагрузки на них
Длина сварочных швов и классов арматуры находятся в прямой зависимости. В работе используются электроды с сечением 4—5 мм. Длина нахлеста при проведении сварочных работ — менее 10 диаметров используемых прутьев, что соответствует требованиям регламентирующих ГОСТов 14098 и 10922.
Длина нахлеста при проведении сварочных работ — менее 10 диаметров используемых прутьев, что соответствует требованиям регламентирующих ГОСТов 14098 и 10922.
Стыковка арматуры методом вязки
Это самый простой способ обеспечить надежную конструкцию из арматурных прутьев. Для этой работы используется самый популярный класс стержней, а именно, А400 AIII. Соединение арматуры внахлест без сварки выполняется посредством вязальной проволоки. Для этого два прутка приставляются друг к другу и обвязываются в нескольких местах проволокой. Как говорилось выше, согласно СНиП, есть 3 варианта фиксации арматурных прутьев вязкой. Фиксация прямыми концами периодического профиля, фиксация с прямыми концами поперечного типа, а также пользуясь деталями с загибами на концах.
Выполнять соединение прутьев арматуры внахлест абы как нельзя. Существует ряд требований к этим соединениям, чтобы они не стали слабым местом всей конструкции. И дело не только в длине нахлеста, но и других моментах.
Типы соединения
В действующих строительных нормах и правилах (СНиП) подробно описывается крепление арматуры всеми существующими в настоящее время способами. На сегодняшний день известны такие методы состыковки арматурных прутьев, как:
- Стыки внахлест, выполненные без сварки:
- нахлест при стыковке с помощью изогнутых деталей (петлей, лапок, крюков).
- нахлест в соединениях прямых прутьев арматуры с поперечной фиксацией;
- нахлест прямых концов прутьев.
- Механические и сварные типы соединений встык:
- с использованием сварочных аппаратов;
- при помощи профессиональных механических агрегатов.
Нахлестом рекомендовано соединять арматуру сечением не более 40 миллиметров
В требованиях СНиП сказано о том, что в бетонном основании необходимо устанавливать как минимум 2 неразрывных арматурных каркаса. Они выполняются фиксированием армирующих прутьев внахлест. Вариант сплетения прутьев внахлест популярен в частном строительстве.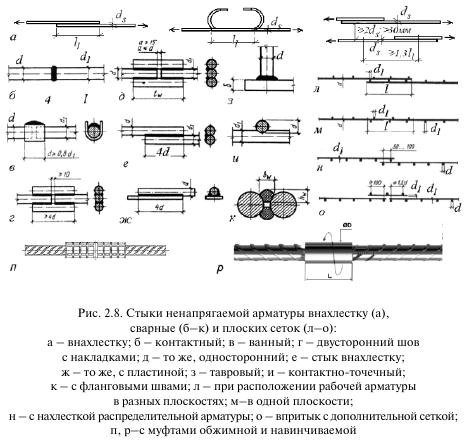
И этому есть объяснение — такой способ доступен, а необходимые материалы имеют невысокую стоимость. Состыковать нахлест стержней арматуры без применения сварки можно с использованием вязальной проволоки.Промышленное строительство чаще использует второй вариант соединения арматурных прутьев.
Строительными нормами допускается во время соединения арматуры внахлест применение прутьев разных сечений (диаметров). Но они не должны превышать 40 мм из-за отсутствия технических данных, подтвержденных исследованиями. В тех местах, где нагрузки максимальны, запрещается фиксация внахлест как при вязке, так и в случае использования сварки.
Точечная контактная сварка
Контактная сварка – это самый распространённый и современный метод соединения. Суть его заключается в пропускании тока по цепи (стержни так же являются ее частью). При этом в месте контакта происходит нагревание, деформация и последующее сдавливание.
Суть его заключается в пропускании тока по цепи (стержни так же являются ее частью). При этом в месте контакта происходит нагревание, деформация и последующее сдавливание.
Точечное соединение чаще всего используется для создания сеток из арматуры малого диаметра. Также данный вид подойдет при ремонте оборудования и деталей.
Существует два способа выполнения шва. В первом случае металл нагревают, а во втором – на участке соединения его доводят до жидкого состояния (формируется литое ядро).
Для самого процесса точечной сварки необходимо дорогостоящие оборудование, которое способно дать высокие показатели тока. Его ключевым узлом выступает трансформатор.
Процесс сварки выполняется в два основных этапа:
- Прутки укладываются и фиксируются в определенном положении между электродами.
 Они должны быть плотно прижаты и статичны.
Они должны быть плотно прижаты и статичны. - Подается ток, и металл нагревается до пластического состояния. За счет сжатия формируется прочное соединение.
Как стыковать арматуру в колоннах
Архив рассылки «Непрошеные советы» для начинающих проектировщиков. Выпуск № 8.
Доброе утро!
Как и обещала, в этом выпуске я расскажу о стыковке рабочей арматуры в колоннах.
Сначала хочу поговорить о стыковке внахлестку. Если вы выбрали именно этот способ, то нужно всегда помнить, что увязывать расположение арматуры должен проектировщик, а не строители. Если в проекте не будет оговорено положение и форма выпусков арматуры, их отогнут случайным образом или не отогнут вовсе. А после бетонирования колонны гнуть выпуски без нагрева арматуры (а это запрещено нормами) невозможно. В итоге, кое-как торчащая арматура может, во-первых, помешать укладке арматуры балок (если таковые имеются), а во-вторых, и это хуже, помешать нормально установить арматуру выше стоящей колонны.
Как нужно показывать изгибаемый стержень на чертеже? Например, у нас колонна высотой 2900 мм, толщина перекрытия 180 мм, арматура класса А400С диаметром 16 мм, бетон класса В25.
Объясню по пунктам:
- Чтобы в вышестоящей колонне арматура стала на то же место, что и в нижестоящей (особенно угловая), нужно изогнуть выпуск минимум на 20 мм. Не на 16 мм, обратите внимание! Т.к. 16 мм – это номинальный диаметр, по факту он больше за счет выступов на арматуре. Если гнуть больше, чем на 20мм, с запасом, тогда стержни будет сложно подвязать друг к другу.
- 2920 мм + 160 мм = сумма высоты этажа и толщины перекрытия, в данном случае место гиба стержня находится в толще перекрытия. 1300 мм – это длина нахлестки арматуры для стержня диаметром 16 мм в бетоне класса В25 (в данном случае, это одна длина нахлестки – об проблеме выбора длины нахлестки я писала в прошлом выпуске).
- R=48 – это радиус загиба стержня. Рабочую арматуру строители обязаны гнуть с помощью специальных устройств, без нагрева стержней, обеспечивая при гибке требуемый радиус загиба, который проектировщик должен заказать в проекте.
 Если на этом не делать ударения в проекте, то строители точно сами инициативу проявлять не будут. Для арматуры класса А400С (А III) минимальный радиус загиба стержней можно узнать из Руководства по проектированию бетонных и железобетонных конструкций из тяжелого бетона (таблица 33): для стержней диаметром меньше 20 мм радиус загиба равен 3d, для диаметра 20 мм и более – 4d, где d – диаметр стержня.
Если на этом не делать ударения в проекте, то строители точно сами инициативу проявлять не будут. Для арматуры класса А400С (А III) минимальный радиус загиба стержней можно узнать из Руководства по проектированию бетонных и железобетонных конструкций из тяжелого бетона (таблица 33): для стержней диаметром меньше 20 мм радиус загиба равен 3d, для диаметра 20 мм и более – 4d, где d – диаметр стержня.
Иногда, особенно при наличии балок перекрытия, необходимо указывать в проекте не только форму стержня, но и положение выпусков – как они должны быть повернуты, чтобы разминуться с верхней арматурой балки. Сейчас объясню на примере. Есть у нас колонна, армируемая 8 стержнями (на рисунке – голубым цветом) и балка с нижней арматурой (желтым) из трех стержней (от колонны до колонны) и верхней арматурой (синим) из трех стержней над колонной – вся арматура диаметром 16 мм. Зеленым показана рабочая арматура колонны следующего этажа.
Теперь посмотрим, что же будет, если мы не дадим информацию с сечения 3-3 на чертеже? Для нижней арматуры ситуация особо не изменится (см.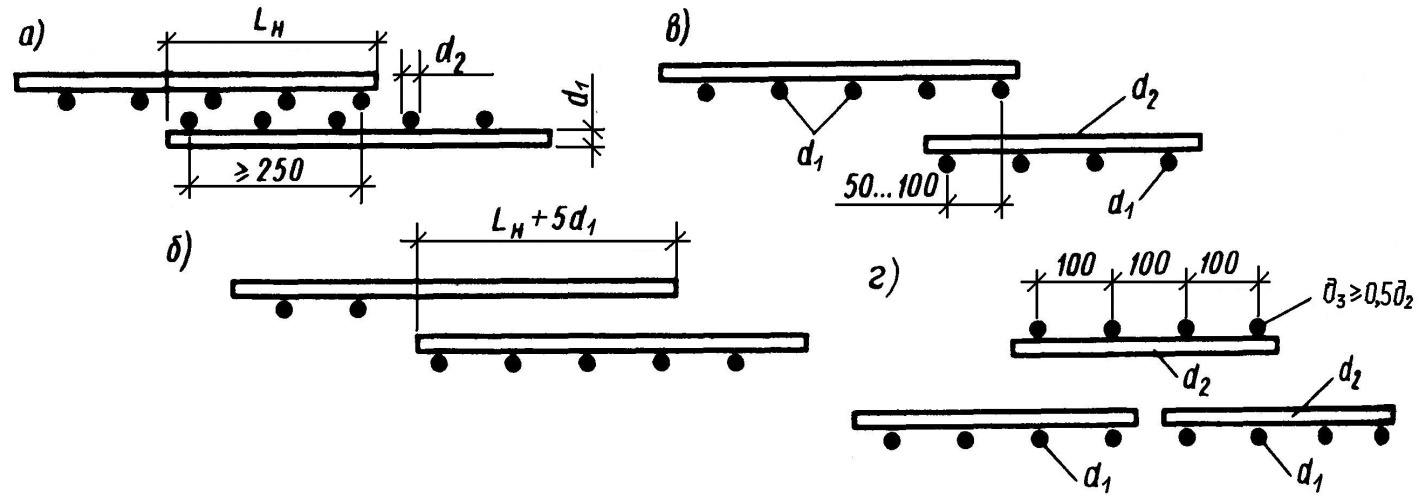 сечение 1-1). Стержни над колонной мы все равно прерываем – их можно подогнуть и развернуть как угодно, лишь бы в бетоне были. А вот верхней арматуре можно чувствительно навредить. Допустим, выпуски не будут развернуты, как следует, и займут место верхней арматуры балки. Куда ей деваться? Разорвать нельзя – это верхняя арматура, ей не хватит длины анкеровки. Отодвинуть от края? Тогда защитный слой для рабочей арматуры будет больше допустимого, да и в углах хомутов арматуры не окажется – плохо.
сечение 1-1). Стержни над колонной мы все равно прерываем – их можно подогнуть и развернуть как угодно, лишь бы в бетоне были. А вот верхней арматуре можно чувствительно навредить. Допустим, выпуски не будут развернуты, как следует, и займут место верхней арматуры балки. Куда ей деваться? Разорвать нельзя – это верхняя арматура, ей не хватит длины анкеровки. Отодвинуть от края? Тогда защитный слой для рабочей арматуры будет больше допустимого, да и в углах хомутов арматуры не окажется – плохо.
А если не дать вообще информацию о том, что арматуру колонны нужно гнуть, и как именно нужно гнуть? Тогда «зеленым» стержням колонны следующего этажа вообще деваться некуда будет.
Вывод: очень важно дать в проекте информацию о форме стержней и их положении в пространстве.
Теперь пару слов скажу о стыковке арматуры сваркой. Оптимальный способ сварки стержней колонны – это сварка с накладками (ГОСТ 14098-91-С21-Рн, или ДСТУ Б В.2.6-169:2011 – сама я этот ДСТУ в глаза не видела, но наш техотдел клянется, что от ГОСТ отличается лишь название).
Минимум, который вы должны учесть в проекте – это указание ссылки на ГОСТ 14098-91-С21-Рн, а то строители приварят прихватками и никто не будет виноват, кроме проектировщика. В идеале необходимо сделать узел стыковки арматуры, заказать накладки, указать длину сварных швов и указать положение накладок относительно граней колонны. Насчет последнего поясню, ситуация подобна с положением выпусков арматуры. Особенно важно указать, где должны быть накладки, для угловых стержней колонн. Иначе строители приварят так, что защитного слоя бетона до арматуры не останется – особенно при больших диаметрах арматуры.
Еще желательно указывать о стыковке сварных швов вразбежку – чтобы в сечении было не более 50% сварных швов.
Еще для общего развития советую найти и почитать СТО 02495307-001-2007 «Сварные соединения арматурных стержней в монолитных железобетонных колоннах зданий и сооружений». Я понимаю, что это стандарт организации и ссылаться на него не корректно, но в нем много хороших решений и отличных идей, опробованных на практике, например вот таких:
Надеюсь, эта информация была полезной для вас! Интересных вам проектов!
С уважением, Ирина.
Правильное армирование углов ленточного фундамента
Содержание статьи
- 1 Правила армирования углов
- 2 Схемы армирования углов
- 2.1 Схема внахлёст (лапки)
- 2.2 Хомут Г-образной формы
- 2.3 Хомут П-образной формы
- 2.4 Тупой угол
- 3 Армирование примыканий
- 3.1 Соединение внахлёст
- 3.2 Хомут Г-образной формы
- 4 Типичные ошибки
Углы и примыкания ленточного фундамента являются местами концентрации разнонаправленных напряжений. Неправильная стыковка продольной рабочей арматуры на участках примыканий и по углам может привести к появлению поперечных трещин, расслоений и отколов в этих проблемных зонах. Правильное армирование ленточного фундамента обеспечивает сопротивляемость железобетонной конструкции силам сжатия и растяжения на всех его участках.
Рис.1. Нагрузки на угол фундамента.
Правила армирования углов
Общие правила применения арматуры при строительстве ленточных фундаментов изложены в СП 50-101-2004.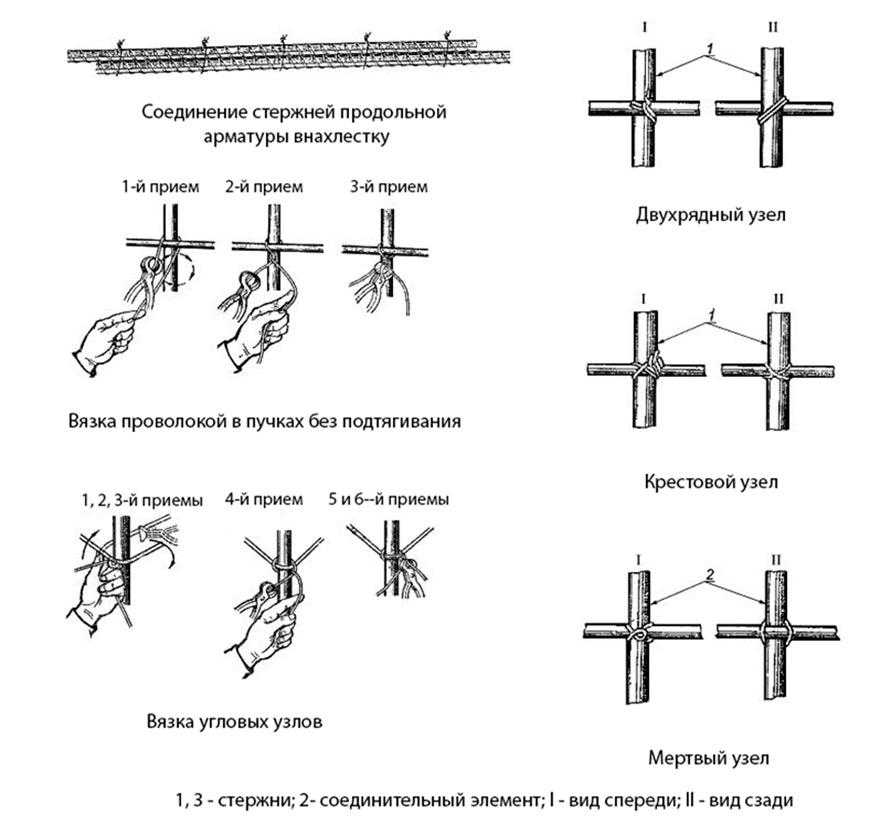 В пункте 8.9 этого документа указано, что фундаменты стен должны объединяться в систему перекрёстных лент и иметь между собой жёсткую связку. О способах жёсткого соединения арматуры говорится в СП 52-101-2003. В пункте 8.3.26 перечислены все допустимые способы таких соединений:
В пункте 8.9 этого документа указано, что фундаменты стен должны объединяться в систему перекрёстных лент и иметь между собой жёсткую связку. О способах жёсткого соединения арматуры говорится в СП 52-101-2003. В пункте 8.3.26 перечислены все допустимые способы таких соединений:
1. Стыковка арматуры без сварки, внахлёст. Допускаются следующие способы анкеровки в районе нахлёстки: с прямыми концами рифлёной арматуры, с приваркой поперечных стержней, с загибами на концах в форме крюков, или петель.
2. Сварка арматуры.
3. Применение механических устройств, или резьбовых муфт.
Жёсткость соединения арматуры на углах, или примыканиях может быть обеспечена только этими способами. Соединения при помощи вязки перекрестий при армировании углов ленточного фундамента не допускаются. В этом случае происходит угловой разрыв арматурного каркаса и потеря его целостности. Для усиления угловых арматурных стыков можно применять П- и Г-образные элементы, изготовленные из арматурных прутьев, применяемых для устройства продольной (рабочей) арматуры. Вертикальные и поперечные хомуты в области угловых и примыкающих анкеровок устанавливаются в 2 раза чаще, чем в остальных частях ленточного фундамента. Оптимальное расстояние между хомутами в зонах примыканий и углов определяется как половина от ¾ высоты ленты. Не рекомендуется делать это расстояние более 25 см. Для равномерного распределения нагрузок на углах ленты, а также в области примыканий, делается жёсткая связка внутренней и внешней продольной арматуры.
Схемы армирования углов
Для формирования единой жёсткой пространственной рамы ленточного фундамента применяют следующие схемы угловых и примыкающих соединений продольной арматуры:
1. Жёсткое угловое соединение арматуры внахлёст и «лапкой».
2. Армирование угловой зоны при помощи хомута Г-образной формы.
3. Схема армирования угла при помощи П-образного хомута.
4. Армирование зоны примыкания при помощи соединения внахлёст.
5. Схема армирования примыкающей зоны при помощи хомута Г-образной формы.
6. Армирование области примыкания при помощи хомута П-образной формы.
7. Армирование тупых углов при помощи жёсткого соединения внахлёст.
Любая из вышеперечисленных схем предусматривает жёсткое соединение внутренней и внешней продольной арматуры.
Схема внахлёст (лапки)
1. Жесткость углового соединения внешней горизонтальной арматуры обеспечивается внахлёст при помощи сгиба одного из свободных концов (1-2).
2. Привязка внутренней горизонтальной арматуры (7) к внешней горизонтальной арматуре (2) осуществляется внахлёст.
3. Привязка внутренней горизонтальной арматуры (3) к внешней связке (1-2) производится при помощи соединения «лапка».
4. Шаг угловой поперечной арматуры (5) и вертикальной арматуры (4) рассчитывается по формуле 3/8 высоты ленточного фундамента.
5. Длина «лапки» составляет 35-50 диаметров продольной арматуры.
Рис. 2. Схема армирования угла внахлёст.
Хомут Г-образной формы
1. Жесткость соединения внешней продольной арматуры (1) в угловой зоне обеспечивает Г-образный хомут (6).
2. Внутренняя продольная арматура (2) жестко скрепляется с внешней продольной арматурой (1) внахлёст.
3. Шаг поперечной арматуры (L) составляет не более ¾ высоты ленты фундамента.
4. Внутреннюю и внешнюю продольную арматуру соединяет дополнительная поперечная арматура (5).
5. Длина соединения внахлёст составляет 50 диаметров горизонтальной арматуры.
Рис. 3. Схема армирования угла г-образным хомутом.
Хомут П-образной формы
1. При использовании П-образных хомутов (5) угловое соединение внешней и внутренней горизонтальной арматуры ленточного фундамента (1) получает жёсткую сцепку наподобие замка.
2. В анкеровке П-образных хомутов участвует вертикальная (2), поперечная (3) и дополнительная поперечная (4) арматура.
Рис. 4. Схема армирования углов п-образным хомутом.
Тупой угол
1. Для надёжного соединения арматурного каркаса при повороте ленточного фундамента под тупым углом (1) используется схема жёсткого соединения внахлёст свободных концов внутренней горизонтальной арматуры (4) с внешней горизонтальной арматурой (5).
2. Вертикальную (2) и горизонтальную (3) арматуру в зоне соединения внахлёст следует устанавливать в 2 раза чаще, чем на ровных участках ленты.
3. Длина соединения внахлёст должна быть не меньше 50 диаметров продольной арматуры.
Рис. 8. Схема армирование тупого угла.
Армирование примыканий
Соединение внахлёст
1. Соединение горизонтальной арматуры (2) примыкающего элемента ленточного фундамента внахлёст осуществляется только к внешней горизонтальной арматуре (1).
2. Шаг поперечной (4), дополнительной поперечной (5) и вертикальной арматуры в зоне примыкания должен быть не менее 3/8 от высоты ленты фундамента.
3. Размеры соединения внахлёст составляют 50 диаметров рабочей арматуры.
Рис.5. Схема армирования примыкания внахлёст.
Хомут Г-образной формы
1. При использовании Г-образного хомута (6) для армирования зоны примыкания горизонтальная арматура примыкающей части и внешняя горизонтальная арматура (1) соединяются с уголком внахлёст.
2. Длина соединения внахлёст (2) составляет 50 диаметров рабочей арматуры.
3. Шаг вертикальной (3) и поперечной арматуры (4) в зоне примыкания уменьшается в два раза при помощи дополнительной поперечной арматуры (5).
Рис. 6. Схема армирования примыкания хомутом г-образной формы.
Хомут П-образной формы
1. Хомут П-образной формы (6) обеспечивает дополнительную жёсткую привязку внахлёст горизонтальной арматуры примыкающего элемента ленточного фундамента (3) к внешней горизонтальной арматуре (1).
2. Длина соединения внахлёст (2) может составлять 35-50 диаметров горизонтальной арматуры.
3. Минимально допустимая длина П-образного хомута должна равняться двойной ширине ленточного фундамента.
Рис. 7. Схема армирования примыкания ленточного фундамента хомутом г-образной формы.
Рекомендуем: Пример расчета диаметра арматуры для ленточного фундамента.
Типичные ошибки
Все способы угловых и примыкающих соединений арматуры направлены на сохранение целостности арматурного каркаса, независимо от его конфигурации. Прочность ленточного фундамента зависит от правильной анкеровки концевых элементов продольной арматуры. К неправильному армированию углов ленточного фундамента приводят следующие схемы:
1. Армирование угловых зон ленточного фундамента арматурными перекрестиями с вязкой стержней продольной арматуры под прямыми углами.
2. Установка в угловых и примыкающих зонах гнутой продольной арматуры без анкеровки.
Эти ошибки являются самыми распространёнными и могут привести к разрушению фундамента в местах угловых соединений и примыканий.
Угловые и примыкающие соединения, выполненные методом вязки перекрестий стержней продольной арматуры
Типичной ошибкой армирования углов и примыканий являются соединения продольной арматуры методом вязки перекрестий. Такое арматурное соединение без надлежащей анкеровки стержней может привести к разрушению бетонного монолита из-за разнонаправленных нагрузок, возникающих по углам ленточного фундамента.
Рис. 9. Частая ошибка при армировании углов
Применение гнутой продольной арматуры для армирования угловых соединений и примыканий
1. Угловые соединения без связки внутренней и внешней продольной арматуры (1) не обеспечивают жесткой стержневой фиксации.
2. Разрушение фундамента может происходить не только из-за образования поперечных трещин, но и из-за отслаивания внутренних углов.
Рис. 10. Ещё один пример неправильного армирования углов
Обязательно прочитайте: Можно ли армировать ленточный фундамент стеклопластиковой арматурой, если собираетесь ее использовать.
Чтобы не допустить появление на углах и примыканиях ленточного фундамента образование трещин, отколов и расслоений, необходимо правильно связать концевые стержни продольной арматуры и выполнить их надёжную анкеровку. Правильное армирование углов ленточного фундамента – залог надёжности и долговечности здания.
часто задаваемых вопросов о фибробетоне | Ярко Поставка | Янгсвилл, Северная Каролина — Фуки Варина, Северная Каролина — Маклинсвилл, Северная Каролина
Поделиться
«Часто задаваемые вопросы по фибробетону»
- Поделись этим:
Поделиться через фейсбук
Пин на Pinterest
Твитнуть в Твиттере
Часто задаваемые вопросы из фибробетона
ЗАЧЕМ ИСПОЛЬЗОВАТЬ WWM ?
Вторичное ненесущее армирование, такое как проволочные маты, не препятствует возникновению трещин, но традиционно используется для скрепления бетона после того, как он растрескается. Синтетические волокна в первую очередь доказали свою способность препятствовать возникновению ранних трещин пластической усадки, а правильно подобранное макроволокно также может влиять на поведение после образования трещин.
Могут ли моноволокна заменить рулонную сварную проволочную ткань (WWF) в бетоне?
НЕТ. За исключением Джаркомеша Типа 2. Некоторые производители волокна рекомендуют однонитевое моноволокно вместо сетки из катаной проволоки в качестве вторичного армирования. Исследования показали, что, хотя волокна моноволокна действительно уменьшают пластическую усадку в начале жизни бетона, их преимущества ограничены, когда бетон трескается. Jarcomesh Type 2 прошел оба критерия тестирования ICC ES AC 32, чтобы заменить WWF.
Могут ли фибриллированные волокна заменить проволочную сетку в бетоне?
ДА. Если проволочная сетка носит неструктурный характер, то фибриллированное (сетчатое) полипропиленовое волокно в минимальной дозировке 1,5 фунта. на кубический ярд (0,9 кг на кубический метр) могут адекватно заменить проволочную сетку в качестве вторичной арматуры, если они соответствуют требованиям ICC минимум 50 фунтов на квадратный дюйм. Jarcomesh Type 2 с весом 2/3 фунта на ярд также может заменить проволочную сетку с давлением 60 фунтов на квадратный дюйм и пройти испытание на удар.
Уменьшают ли синтетические волокна растрескивание бетона?
ДА. Использование синтетических волокон в дозировке, рекомендованной производителем на кубический ярд, может уменьшить растрескивание бетона при пластической усадке. Рекомендуется проконсультироваться с поставщиком волокна и запросить результаты испытаний, и вы обнаружите, что Jarcomesh Type 2 превосходит все другие волокна.
Влияет ли использование фибры на прочность бетона на сжатие?
Использование малых или больших объемов синтетических волокон не предназначено для повышения исходной прочности бетона. Использование волокон заметно не увеличивает или уменьшает прочность на сжатие. Однако было показано, что высокие дозы или макро/структурные синтетические волокна резко меняют характер растрескивания и разрушения бетона, способствуя очень пластичному типу разрушения.
Требует ли использование волокна изменения состава смеси?
ДА И НЕТ. Когда волокна используются в стандартных дозировках и нормах внесения, нет необходимости в изменении состава смеси. Однако при резком увеличении объемного расхода волокна могут потребоваться некоторые изменения в составе смеси. Пожалуйста, свяжитесь с нами для получения помощи относительно дизайна смеси и дозировки волокна.
Устраняет ли использование фибры необходимость применения надлежащих методов бетонирования?
НЕТ. Использование любого синтетического волокна не отменяет необходимости использования бетона. Как и в случае с любым бетоном, важно следовать надлежащим рекомендуемым в отрасли методам смешивания, укладки, соединения и отверждения бетона.
Почему Jarco Supply предлагает различные типы армирования волокнами?
В ходе исследований и разработок было получено несколько марок армирующих волокон для различных областей применения и уровней производительности. Каждый сорт волокна предлагает выдающиеся эксплуатационные характеристики при соответствующем применении.
В чем разница между моноволокном и фибриллированным волокном?
Как следует из названия, моноволокна представляют собой одножильные волокна, по форме напоминающие леску. Фибриллированные волокна деформированы или имеют неправильную форму и расширяются в виде сети, похожей на рыболовную сеть.
Какой тип клетчатки и дозировку рекомендует Jarco Поставка ?
Jarco Supply предлагает ряд синтетических волокон, используемых в различных дозировках, чтобы соответствовать требованиям проекта или владельца. Jarco Supply рекомендует следующие эксплуатационные характеристики:
1. Для предотвращения образования трещин при пластической усадке на ранних стадиях жизни бетона: 1 мешок на ярд Jarcomesh, тип 1 большинство применений: 1 мешок на ярд Jarcomesh Type 2
на ярд Jarcomesh Type 3:
Обратитесь к представителю Jarco по снабжению, чтобы узнать расчетную дозу для каждого применения.
Можно ли перекачивать волокна Jarcomesh ?
Да. Армирование волокном стало желательной строительной практикой для широкого спектра бетонных проектов. Простота добавления и равномерное распределение дают волокнам явные преимущества на строительной площадке по сравнению с неструктурной проволочной сеткой. Эти преимущества еще более ценны в проектах, где бетон подается насосом. Использование встроенного волокнистого армирования устраняет проблемы с проволочной сеткой, с которыми сталкиваются рабочие на насосных линиях, и предоставляет наладчику свободное поле для работы. Вместо того, чтобы поднимать рулоны сетки на проекты настила верхнего уровня, бетон, армированный Jarcomesh, можно просто закачивать на место, что обеспечивает значительную экономию времени и труда для проекта. Хотя волокна имеют тенденцию изменять «внешний вид» бетона, операторы насосов обычно замечают, что для фибробетона требуется более стабильное и немного более низкое давление насоса.
Можно ли использовать волокна Jarcomesh в сборных изделиях?
Да. По определению сборный железобетонный элемент — это просто элемент, который «отлит перед» — тот, который отлит и отвержден в форме, отличной от его конечного положения. Это бетонное изделие может включать в себя широкий спектр элементов: камни для патио, брызговики, ступени, септиктенки, архитектурные фасадные панели, разделительные барьеры, железнодорожные шпалы, склепы, хозяйственные ящики, мостовые балки, ступенчатые кольца, трубы, пустотелые конструкции. стержневые плиты, люки и столбы забора, а также сотни различных декоративных декоративных элементов. Для производителя сборных железобетонных изделий очень важно найти методы повышения ударной вязкости и ранней прочности своих бетонных изделий, чтобы уменьшить количество отходов, свести к минимуму повторные вызовы и возвраты, а также способствовать долговечности изделия. Если сборщики сборных железобетонных изделий могут снимать опалубку и перемещать «зеленые» продукты в зону затвердевания без поломки, то армирование волокном, очевидно, выполняет свои первоначальные эксплуатационные требования. Кроме того, производители сборных железобетонных изделий замечают меньше поломок, сколов и растрескивания во время обработки, доставки и размещения своей продукции благодаря уникальному трехмерному покрытию волокна Jarcomesh. Использование более высоких доз макроволокон позволяет производителю сборного железобетона заменить обычную сталь более высокого уровня. Обратитесь в Jarco Supply для получения технической помощи.
Можно ли использовать Jarcomesh для торкретирования?
Да. Термин «торкретбетон» обычно используется для описания бетона или строительного раствора, который укладывается или распыляется с высокой скоростью на заданную поверхность с помощью сжатого воздуха. Армирование, используемое в типичных применениях торкретбетона, должно обеспечивать устойчивость к нагрузкам на сдвиг, изгиб и изгиб, которые могут возникнуть в результате движения грунта или горных пород или местного гидростатического давления. Размещение проволочной сетки на типичных неровных поверхностях торкретбетона является обременительным и дорогостоящим с точки зрения труда. Синтетические волокна могут использоваться в качестве альтернативных материалов, обеспечивающих необходимый индекс прочности и требуемые уровни остаточной прочности, без хлопот и трудозатрат, связанных с сеткой.
Можно ли использовать волокна Jarcomesh для приподнятых плит?
Да. Существует ряд терминов, используемых для описания систем с приподнятыми плитами, таких как плита на металлическом настиле и композитный настил. Элементами этой системы являются металлический настил, бетон на портландцементе и, в большинстве случаев, какая-либо арматура. Металлический настил можно разделить на три категории: структурный (композитный), опалубочный и кровельный. Первым шагом является выбор подходящего металлического настила для применения. Как правило, в большинстве многоэтажных конструкций используется составной (несущий) настил пола, при этом настил выступает в качестве основного или положительного армирования. И наоборот, в системе некомпозитного настила металлический настил используется только в качестве формы, основная или положительная арматура которой будет встроена в бетонную плиту. В системе составного стального настила сварная проволочная сетка иногда используется в качестве температурного или вторичного армирования. Расчет сварной проволочной сетки для температурного и усадочного армирования согласно Институту стальных настилов составляет 0,00075 площади бетона над настилом, однако SDI продолжает утверждать, что «если используется сварная проволочная сетка с площадью стали, указанной выше формуле, как правило, будет недостаточно полного отрицательного подкрепления». Это соображение позволяет использовать волокна Jarcomesh Macro в качестве замены сварной проволочной ткани в качестве вторичного армирования. Эти волокна обеспечивают однородное трехмерное вторичное армирование, которое превосходит любую другую форму температурного/вторичного армирования, а также являются более безопасными и экономичными в использовании. В любых приложениях выше класса следует проконсультироваться с Jarco Supply для получения помощи в расчете армирования.
Можно ли использовать волокна Jarcomesh в начинках или покрытиях?
Да. Верхний слой определяется как слой бетона или раствора, редко тоньше 1 дюйма (25 мм), уложенный на изношенную или потрескавшуюся поверхность бетонной плиты и обычно приклеенный к ней. Наложение обычно предназначено либо для восстановления, либо для улучшения функции предыдущей поверхности. Точно так же верхнее покрытие также определяется как слой бетона или раствора, уложенный для формирования поверхности пола на бетонном основании, но не обязательно связанный с существующей плитой. Хотя износ старой поверхности или сильное растрескивание старой плиты чаще всего является причиной нанесения верхнего слоя, другие причины могут включать отсутствие ровности пола, неправильную высоту или плоскость, недостаточное сопротивление скольжению или скольжению или отсутствие износостойкости. . Независимо от причин, накладки и накладки на плиты могут стать экономически эффективным методом восстановления существующей плиты до рабочего состояния без затрат на демонтаж и замену. В дополнение к обычным трудностям размещения сетки в плоских изделиях существуют дополнительные сложности, связанные с размещением накладок и накладок. Естественно, стальная проволочная сетка требует достаточного покрытия в бетоне (обычно не менее 2 дюймов или 5 см), чтобы предотвратить отслаивание, связанное с коррозией, и неприглядные линии сетки. Очевидно, что такое покрытие становится невозможным в тонких бетонных покрытиях. , укладка проволочной сетки становится одинаково затруднительной без нарушения или повреждения разрывного слоя или защитного покрытия. Одним из наиболее важных минусов сетки является отсутствие равномерного армирующего покрытия.Сетка явно расположена в одной плоскости только в этих тонкие приложения, требующие армирования для устранения проблем, вызванных однонаправленным вытеканием, дифференциальной усадкой и скручиванием.
Когда лучше всего добавлять волокна Jarcomesh в бетон?
Продукты Jarcomesh должны быть добавлены в систему смешивания бетона на бетонном заводе для лучшего распределения. Следуйте стандартным рекомендациям производителей обычных смесителей и ASTM C-94. Время перемешивания должно составлять не менее четырех-пяти минут на одну загрузку при нормальной скорости перемешивания. Завод периодического действия будет наиболее экономичным и безопасным местом для добавления волокон. Обычно не рекомендуется вводить волокна в смеситель в качестве первого ингредиента, а добавлять их вместе с другими ингредиентами или в конце последовательности добавления.
Не вызовет ли проблемы добавление Jarcomesh волокон на рабочей площадке?
Волокна можно добавлять в грузовики для готовых смесей на стройплощадке, хотя рекомендуется добавлять их на заводе по производству шихты для оптимального смешивания и распределения. Если волокна добавляются на месте, следует проявлять особую осторожность, чтобы обеспечить достаточное время перемешивания. Выждите не менее 4-5 минут для перемешивания при скорости барабанного перемешивания после добавления последнего мешка с продуктом.
Совместимы ли волокна Jarcomesh с жидкими добавками?
Синтетические волокна не влияют на воздухововлечение, суперпластификаторы или понизители воды. Если возможно, синтетические волокна следует добавлять перед любыми жидкими добавками, чтобы в полной мере использовать сдвиг и трение смеси при смешивании для оптимизации распределения.
Будут ли волокна Jarcomesh мешать нанесению лазерной стяжки или затирочной машины?
НЕТ, вибрация стяжки с лазерным наведением выносит цементную пасту на поверхность и покрывает почти все открытые волокна. Те, что не покрыты, будут сожжены любой затирочной машиной. Возможность замены обычных стальных матов синтетическими волокнами большого объема значительно упрощает укладку и отделку лазерной стяжки.
Какой процесс следует использовать при нанесении щетки?
Использование щетки с жесткой щетиной только в одном направлении поможет совместить поверхностные волокна с гребнями текстуры, что сделает их значительно менее заметными.
Мешают ли волокна адгезии герметиков или напольных покрытий?
Поверхностные волокна не вступают в реакцию с герметиком и/или не мешают ковровому покрытию, плитке и т. д. При необходимости можно использовать тепловую горелку для удаления любых волокон, которые могут вызывать беспокойство.
Какое влияние оказывает фибра в бетоне на осадку?
Из-за своей трехмерной когезивной природы бетон, армированный волокнами, выглядит менее пригодным для обработки, чем обычный бетон. В действительности, визуальная осадка может немного уменьшиться, но текучесть остается почти такой же. Осторожность; никогда не допускайте добавления воды на рабочем месте, чтобы вернуть потерю осадки. При необходимости рекомендуется использовать суперпластификатор, чтобы увеличить осадку.
Признаны ли волокна Jarcomesh органами национального законодательства США?
Да. Компания Jarcomesh протестировала все свои волокна на соответствие всем нормам и стандартам, используемым ICC. Все национальные строительные нормы и правила, такие как Единые строительные нормы и правила (I.C.B.O. — Международная конференция строительных норм и правил), Стандартные строительные нормы и правила (S.B.C.C.I. — Международный конгресс южных строительных норм), Основные строительные нормы и правила (B.O.C.A. — Администраторы кодекса строительных служащих), и Жилищный кодекс для одной и двух семей (C.A.B.O. — Совет американских строительных чиновников). Эти три кодекса теперь объединены в I.C.C. Международный совет по нормам и правилам), по которому тестируются все продукты Jarcomesh.
Все ли стальные волокна одинаковы?
Нет – Характеристики стального волокна зависят от дозировки, предела прочности на растяжение, соотношения сторон и анкеровки. Совместное влияние этих четырех факторов на бетон определяют путем испытаний в соответствии с ASTM C1609 (Стандартный метод испытаний на изгиб фибробетона с использованием балки с нагрузкой в третьей точке). По результатам испытаний можно определить среднюю эквивалентную прочность на изгиб (EFS) железобетона. EFS – это проверенная прочность железобетона после появления трещин.
Какое отношение денье и соотношение сторон имеют к волокну?
Денье волокна – это мера массы отдельной пряжи или нити волокна на длине 9000 м. Обычно это используется только при производстве синтетических материалов и используется для процедур ОК/КК. Соотношение сторон волокна — это длина отдельного волокна, деленная на его эквивалентный диаметр (L/d). Этот термин обычно используется только с более крупными волокнами, такими как сталь и макросинтетика, и, хотя конкретное значение не имеет значения, соотношение сторон более 100 иногда может вызывать трудности с укладкой и отделкой.
Почему волокна в бетонных смесях «комкуются»?
Волокна всех типов (стальные, микро- и макросинтетические) могут слипаться в бетоне. Это явление обычно вызывается добавлением фибры в слишком сухие бетонные смеси (подвижность снижается до нуля) или в смеси, в которых недостаточно мелких частиц (цемент, песок, добавки и т. в свою очередь, паста «голодает» систему и снова заставляет спад уменьшаться до нуля. Свободные волокна в пустом барабане могут слипаться, а слишком длинные волокна или волокна с различной геометрией также могут вызывать проблемы. Как всегда, необходимо провести пробное испытание, чтобы убедиться, что смесь соответствует типу волокна и дозировке и что последовательность дозирования не вызовет никаких проблем. При необходимости может быть оправдано использование водопонижающей добавки для поддержания желаемой осадки при укладке.
Можно ли использовать микроволокна с высокой дозировкой вместо макроволокон с низкой дозировкой?
Возможно — Опять же, ключом будет дозировка и предполагаемая функция волокон. Основной функцией микросинтетического волокна является контроль трещин пластической усадки, и исследования показали, что эти волокна не обладают значительной способностью выдерживать нагрузку через трещину. Хотя данные испытаний могут поддерживать использование микроволокна, это может быть не лучшим вариантом. Во-вторых, большие дозы микросинтетики будет сложнее смешивать, так как количество волокон и площадь поверхности волокон будут чрезвычайно велики, что может привести к значительным потерям при оседании.
Все ли макросинтетические волокна одинаковы?
Нет — на рынке представлено несколько различных типов макросинтетики, каждая из которых имеет свои преимущества и преимущества. Помните старую пословицу; «ты получаешь то, за что платишь». Ключом к успешному использованию макросинтетического волокна для замены WWM, арматуры или стальных волокон является дозировка. Для более прочных волокон или волокон с более высоким сцеплением, вероятно, потребуется меньше материала, чем для более слабых волокон или волокон с меньшей связующей способностью. Производитель должен сопровождать значения дозировки информацией об испытаниях. Если вопросы все еще присутствуют, следует провести пробную версию, чтобы убедиться, что желаемая производительность достигнута.
Как вы классифицируете арматуру из стального волокна для бетона?
Стальная фибра определяется в ASTM A820 как куски гладких или деформированных волокон, которые достаточно малы, чтобы быть рассеянными в бетонной смеси случайным образом. В настоящее время существует 5 обозначений стальной фибры в зависимости от продукта или процесса, используемого в качестве исходного материала:
Тип I – холоднотянутая проволока
Тип II – листовой прокат
Тип III – экстракция из расплава
Тип IV — фрезерование
Тип V — модифицированная холоднотянутая проволока
При обсуждении бетона, армированного стальной фиброй, в ACI 360 говорится, что «стальные волокна имеют более высокий модуль упругости и прочность на растяжение, чем окружающий бетон. Кроме того, многие типы стальных волокон деформируются для оптимизации закрепления в бетоне. Эти свойства позволяют стальной фибре перекрывать трещины, возникающие в закаленном состоянии, и перераспределять накопленное напряжение, вызванное приложенными нагрузками и усадкой».
Можно ли перекачивать бетон, армированный стальной фиброй?
Да, но ожидайте потерь от осадки от 1 до 3 дюймов через шланг в зависимости от дозировки стального волокна, температуры окружающей среды и длины шланга. Реагент для снижения содержания воды в среднем диапазоне (MRWR) обычно используется для повышения работоспособности и облегчения потока через насосные линии. В некоторых случаях могут потребоваться редукторы высокого давления (HRWR). Обычно требуется шланг диаметром от 4 до 6 дюймов.
ПРИЛОЖЕНИЯ
Возможные проекты, подходящие для использования фибробетона, перечислены ниже.
Жилые помещения : в том числе подъездные пути, тротуары, строительство бассейнов с торкретированием, подвалы, цветной бетон, фундаменты, дренаж и т. д.
Коммерческие помещения : наружные и внутренние полы, плиты и парковочные площадки, проезжие части и
Склады / Промышленный : легкие и тяжелые нагруженные полы и дороги
Автомагистрали/проезжие дороги/мосты : обычное бетонное покрытие, SCC, белые покрытия, ограждающие рельсы, бордюры и водосточные желоба, водопроницаемый бетон, звукопоглощающие барьеры и т. д.
Порты и аэропорты погрузочные рампы.
Водные пути : плотины, шлюзовые сооружения, облицовка каналов, канавы, ливневые сооружения и т. д.
Горное дело и туннелирование : Сборные сегменты и бетонобетон, которые могут включать облицовку туннелей, шахты, стабилизацию откосов, канализационные работы, и т.д.
Приподнятые настилы : включая коммерческие и промышленные конструкции из композитных металлических настилов и приподнятую опалубку в аэропортах, коммерческих зданиях, торговых центрах и т. д.
Сельское хозяйство : сооружения для ферм и животных, стены, силосы, мощение и т. д.
Сборный железобетон и изделия : архитектурные панели, подъемно-откидные конструкции, стены, ограждения, септиктенки, усыпальницы, жироуловители, банковские хранилища и скульптуры
Другие приложения : включает любые другие приложения, связанные с FRC, не описанные выше.
ТИПЫ ВОЛОКНА
Типы волокон для использования в FRC Приложениях бывают разных размеров, форм, цветов и вкусов.
Стальные волокна : Эти волокна обычно используются для придания бетону повышенной прочности и несущей способности после образования трещин. Как правило свободные или связанные, эти волокна, как правило, изготавливаются из углеродистой или нержавеющей стали и имеют различную геометрическую форму, например, извитые, с загнутыми концами или с другими механическими деформациями для закрепления в бетоне. Типы волокон классифицируются в ACI 544 как типы с I по V и имеют максимальную длину от 1,5 до 3 дюймов (30–80 мм) и могут быть дозированы от 10 до 100 фунтов/ярд (от 6 до 67 кг/м3).
Микросинтетические волокна : Эти волокна обычно используются для защиты и уменьшения растрескивания бетона при пластической усадке. Большинство типов волокон изготавливаются из полипропилена, полиэтилена, полиэстера, нейлона и других синтетических материалов, таких как углерод, арамид и другие акриловые материалы. Эти типы волокон обычно дозируются в малых объемах в диапазоне от 0,03 до 0,2% от объема бетона — от 0,5 до 3,0 фунтов/ярд (от 0,3 до 0,9 кг/м3).
Макросинтетические волокна : Этот новый класс волокон появился за последние 15 лет в качестве подходящей альтернативы стальным волокнам при правильном дозировании. Типичные материалы включают полипропилен и другие полимерные смеси, имеющие те же физические характеристики, что и стальные волокна (длина, форма и т. д.). Эти волокна можно дозировать от 3 до 20 фунтов/ярд (от 1,8 до 12 кг/м3).
Стекловолокно : GFRC (бетон, армированный стекловолокном) преимущественно используется в архитектурных приложениях и модифицированных панельных конструкциях на основе цемента.
Целлюлозные волокна : целлюлозные волокна, изготовленные из продуктов переработки древесной массы, используются аналогично микросинтетическим волокнам для контроля и уменьшения растрескивания при пластической усадке.
Натуральные волокна : Натуральные волокна обычно не используются в коммерческих целях для армированного фиброй бетона, они используются для армирования продуктов на основе цемента в приложениях по всему миру и включают такие материалы, как кокос, сизаль, джут и сахарный тростник. Эти материалы бывают разной длины, геометрии и характеристик материала.
Волокна ПВС : Волокна из поливинилового спирта представляют собой синтетические волокна, которые при использовании в больших объемах могут изменить характеристики бетона при изгибе и сжатии относится к вновь изготовленным или специфицированным материалам, не относящимся к вышеперечисленным категориям.
Смеси стали и микро-/макроволокон : недавней разработкой в области фибробетона, появившейся на рынке, стала комбинация или смешивание стальных и/или макросинтетических волокон с различными типами микроволокон для помогают контролировать растрескивание при пластической усадке (т.е. микросинтетика), в то же время обеспечивая бетону повышенную прочность и несущую способность после образования трещины, достигаемые только при использовании стальных и макросинтетических волокон. Эти волокна обычно дозируются по преобладающим
Прочие волокна и смеси : Комбинации и типы волокон, не классифицированные выше
404
- Правила и ресурсы
- Услуги
- Инновации и технологии
- Новости и события
- О нас и карьере
- Правила и ресурсы
- Правила и руководства
- Нормативные обновления
- Консультации и отчеты
- Государство флага и порта
- Инженерное программное обеспечение
- Технические обзоры
- Портал ABS MyFreedom™
- Базы данных
- Формы
- Услуги
- Одобрение и сертификация
- Аудиты компаний и судов
- Global Marine
- Глобальный оффшор
- Глобальная оффшорная ветроэнергетика
- Глобальное правительство
- Устойчивое развитие и обезуглероживание
- Цифровые решения
- Кибербезопасность
- Морское обучение
- Новости и события
- Комната новостей
- События
- Публикации
- О нас и карьере
- Кто мы
- Безопасность
- Карьера
- Свяжитесь с нами
Свяжитесь с нами
Электронная почта: CSC@eagle. org
Позвоните нам: 1-281-877-6000
Дом
О нас
Группа АБС
Карьера
Условия использования
Юридическая информация/Конфиденциальность
Политики и уведомления
Site Map
ABS MyFreedom™ Portal
Rules and Guides
Services
ISO & IACS Certifications
ABS App On The Go
© Американское бюро судоходства, 2023 г. Все права защищены.
Соединения и соединения США Патенты и патентные заявки (класс 403)
Выборочное соединение ступицы с валом (класс 403/1)
Включая хрупкий компонент (класс 403/2)
Выборочно собранные, например, конвертируемые и т. д. (Класс 403/3)
Селективная ориентация (Класс 403/4)
Надувной соединитель (Класс 403/5)
С отверстием для доступа к разъему или оператору (Класс 403/6)
К резьбовому компоненту (Класс 403/7)
К смещенному компоненту (Класс 403/9)
Затвор или заглушка на расстоянии от соединения (класс 403/10)
С дополнительной защитой, фиксатором сломанных деталей, функцией ремонта, сборки или разборки (класс 403/11)
Удерживающие средства или защитные приспособления функционируют только во время транспортировки, сборки или разборки (класс 403/12)
Позиционные или направляющие средства (класс 403/13)
Использование давления жидкости (класс 403/15)
Средства обратного отвода, например, сепаратор и т.
 д. (класс 403/16)
д. (класс 403/16)Ручка или маховик (Класс 403/17)
Средства зацепления или размещения инструмента или с инструментом (класс 403/19)
Съемная невыпадающая гайка или болт (Класс 403/21)
Средства для стопорения болта или гайки при повороте другого (класс 403/22)
С дополнительным экраном (Класс 403/23)
Конструктивно установлен в различных художественных устройствах (Класс 403/24)
Голова или основание (Класс 403/25)
Муфта с независимым подшипником (Класс 403/26)
С индикатором или средствами контроля (Класс 403/27)
Использование тепловых характеристик, например, расширение или сжатие и т. д. (Класс 403/28)
Промежуточный материал со средним коэффициентом расширения (Класс 403/29)
Элементы с разными коэффициентами расширения (класс 403/30)
С компонентом, реагирующим на давление жидкости (класс 403/31)
Со средствами управления, реагирующими на условия окружающей среды (класс 403/32)
С независимой рукояткой или приводом (Класс 403/33)
С отличной функцией перемещения или направления жидкости или предмета (класс 403/34)
Внешний резервуар (класс 403/35)
Напорный фитинг (Класс 403/36)
Включая порт или резервуар, сообщающийся с соединением (класс 403/37)
Неприлипающий текучий материал на стыке (Класс 403/40)
Со средствами ограничения деформации (Класс 403/41)
Комбинированный (Класс 403/42)
Вращательно соединенные, дифференциально поступательные элементы, например, талреп и т. д. (класс 403/43)
Имеющие средства захвата инструмента или ручку управления (класс 403/44)
Наличие рабочего механизма (Класс 403/45)
Имеющие средства блокировки (Класс 403/46)
Дифференциальный шаг, аналогичная стрелка (класс 403/47)
Привод с внешней резьбой (класс 403/48)
Боковые элементы, соединенные с помощью защелок, например, соединители лесов (класс 403/49)
Гибкая диафрагма или сильфон (класс 403/50)
С дополнительным соединением или набивкой (Класс 403/51)
Шарнирные элементы (Класс 403/52)
Несколько отдельных осей артикуляции (класс 403/53)
Разветвленный (Класс 403/64)
Элемент, повернутый под жестким углом (Класс 403/65)
Включая статическое соединение (класс 403/66)
Включая оператора (класс 403/81)
Дугообразные перемещения хвостовика в дополнительной направляющей (Класс 403/82)
Запираемый в фиксированном положении (Класс 403/83)
Вращающиеся элементы, упруго смещенные в одно положение (Класс 403/111)
Включая ограничительные средства (Класс 403/112)
Резьбовой интерфейс (класс 403/118)
Поворотный (Класс 403/119)
Поворотный (класс 403/164)
Коаксиальные элементы, подпружиненные в осевом направлении (Класс 403/166)
Поперечный стержень к разнесенным поверхностям пластин (Класс 403/167)
Фиксатор использует несколько пластин или примыкает к ним (Класс 403/168)
Разветвленный (Класс 403/169)
Три или более излучающих элемента (класс 403/170)
Многогранник (Класс 403/176)
Параллельные стержни (Класс 403/177)
Копланарный (Класс 403/178)
Соединительный элемент из различных материалов (класс 403/179)
Три или более последовательных соединения, по крайней мере, одно разнородное (Класс 403/180)
Несоосные оси (класс 403/181)
- Разъемное промежуточное соединение (Класс 403/182)
Соединение, включающее несколько слоев одного стержня (класс 403/185)
Боковые стержни, независимо соединенные с поперечной поверхностью (Класс 403/186)
Стержень к стержню к стороне, например, пластина, сторона стержня и т. д. (Класс 403/187)
Стержень из стороны в сторону (Класс 403/188)
Односторонний боковой (Класс 403/189)
Дугообразная или многоплоскостная сторона (класс 403/190)
Промежуточный элемент включает трубчатую часть (Класс 403/192)
Промежуточный элемент полностью проходит через бок (Класс 403/201)
Радиально расположенные элементы, соединенные независимой муфтой (класс 403/202)
Гибкий промежуточный элемент (Класс 403/203)
Взаимодействующее соединение между внутренним элементом и несколькими частями внешнего элемента (класс 403/204)
Жесткое угловое соединение, например, коленчатое или U-образное и т. д. (Класс 403/205)
Изогнутая в осевом направлении или изогнутая часть стержня является соединительным элементом (класс 403/206)
Внутренние поперечные стержни (Класс 403/207)
Соединение с боковой поперечной поверхностью в осевых точках (Класс 403/208)
Обратно изогнутая часть (Класс 403/209)
Переплетенные концы (Класс 403/214)
Конец стержня отклоняется в сторону внутри соединителя (Класс 403/215)
Множественные стержни, искривленные одним компонентом (Класс 403/216)
Три или более членов, соединенных в одном месте (класс 403/217)
Все включено (класс 403/218)
Взаимно контактирующие (класс 403/219)
Гибко связанные жесткие элементы (класс 403/220)
Жесткая втулка охватывает гибкую втулку (класс 403/221)
Гибкая муфта втулочного типа (Класс 403/223)
Соединительный штифт пересекает радиально расположенный эластомер (Класс 403/224)
Эластомер, проложенный между радиально расположенными элементами (класс 403/225)
Муфта со спиральной пружиной (Класс 403/229)
Конец стержня к поперечной стороне элемента (Класс 403/230)
Угловое соединение (Класс 403/231)
Стремя (Класс 403/232.
1)Конструкция, прикрепленная к концу стержня, охватывает боковую часть (класс 403/233)
Включая седло (класс 403/237)
Тип сальника (Класс 403/238)
Стержень крепится к краю отверстия (Класс 403/240)
Стержень к открытому краю стороны, например, закручивающееся соединение (класс 403/241)
Стержень и сторона деформируемо заблокированы (Класс 403/242)
Радиально расположенная прокладка или втулка с боковым зацеплением (Класс 403/243)
Гнездо, образованное формованием или деформацией стороны (класс 403/244)
Неотъемлемый поперечный выступ сбоку, например, шейка и т.
д. (Класс 403/245)Использование противоположных сторон элемента (класс 403/247)
Фланец на стержне, прикрепленный сбоку болтами (Класс 403/262)
Стержень, вставленный в утопленную сторону, например, гнездо или коническое отверстие и т. д. (Класс 403/263)
Использование внутренней стержневой конструкции (класс 403/264)
Формованное соединение (Класс 403/265)
И независимое соединение (Класс 403/266)
Включая механическую блокировку (класс 403/267)
Гнездо или открытая чашка для склеивания (Класс 403/268)
- Член, отлитый на месте (класс 403/269)
Соединение плавлением, например, сварка и т.
д. (Класс 403/270)
Уменьшенная посадка (Класс 403/273)
Член деформирован на месте (Класс 403/274)
Разделительный элемент между ветвями элемента (класс 403/275)
Промежуточный деформирующий элемент контактирует с нижней частью гнезда (Класс 403/276)
Внутренний элемент расширен за счет продольно вставленного элемента (Класс 403/277)
Отдельным деформируемым элементом (Класс 403/278)
Деформация происходит одновременно с действием отдельного, разнофункционального, суставного компонента (Класс 403/279)
Деформирующий компонент вставной секции (Класс 403/280)
Отдельные деформирующие средства остаются при сборке узла (Класс 403/281)
Деформация происходит одновременно со сборкой (Класс 403/282)
Прокалыванием (Класс 403/283)
Разная устойчивость к боковой деформирующей силе (Класс 403/284)
Оба участника деформированы (класс 403/285).
Соединён диверсифицированным соединителем (класс 403/286)
Включая разнесенные, разнообразные соединения (класс 403/287)
Включая отдельную упаковку (класс 403/288)
Раздвоенный конец с подвижными в поперечном направлении противолежащими частями (Класс 403/289)
С отдельными средствами приложения силы (Класс 403/290)
Гибкий элемент является компонентом соединения (Класс 403/291)
Концы стержня соединены вставленной секцией (Класс 403/292)
Внешнее шунтирование (Класс 403/293)
Секция, вставленная сбоку (Класс 403/294)
Уголок (Класс 403/295)
Секция с резьбой на стержне (Класс 403/296)
Расширяемая секция (Класс 403/297)
Рифленая или шлицевая секция (Класс 403/298)
Резьбовой переходник (Класс 403/299)
Отдельный концевой соединитель (класс 403/300)
Различные последовательные соединения (класс 403/301)
Один привод для нескольких соединений (класс 403/303)
Множественные оппозитные розетки (Класс 403/305)
Отдельные винтовые или штыревые соединения (класс 403/306)
Непрерывная резьба (Класс 403/307)
Наличие съемных торцевых крышек или заглушек (Класс 403/308)
Прерванная периферия, например, расщепленная или сегментарная и т.
д. (Класс 403/309)Включая средства расклинивания или эксцентрика элементов (класс 403/314)
Включая вспомогательные средства, блокирующие разъединение соединения (класс 403/315)
Блокатор для фиксатора шарнира в сборе (Класс 403/316)
Блокировщик — защелка или ключ (класс 403/319)
Кулачковый или резьбовой фиксатор (класс 403/320)
Защелка с ручным открыванием (класс 403/321)
Наличие рабочего механизма (Класс 403/322.
1)Поворотный болт (Класс 403/323)
Поперечно скользящий штифт (Класс 403/324)
Пружинный манипулятор (класс 403/325)
Смещенная защелка или защелка (класс 403/326)
Отдельной пружиной (Класс 403/327)
Листовая рессора (Класс 403/329)
Поворотный (Класс 403/330)
Боковая направляющая: удлиненные коллинеарные элементы (класс 403/331)
Кольцевая выемка является соединительным элементом (Класс 403/332)
Конвергентный интерфейс (класс 403/333)
Конический (Класс 403/334)
Соединённые фланцы или буртики (Класс 403/335)
Отдельный фланец или плечо (класс 403/336)
Осевые болты или заклепки (Класс 403/337)
Зажимной (Класс 403/338)
Шарф (Класс 403/339)
Зеркальные изображения (класс 403/340)
Промежуточные элементы с внешней перемычкой (Класс 403/341)
Наперсток: винт или кулачок (класс 403/342)
Винт или кулачок (класс 403/343)
Осевой разъемный или разъемный элемент (Класс 403/344)
Межосевые элементы (класс 403/345)
Скрещенные стержни (Класс 403/346)
Элемент с проушинами, поворотное зацепление (Класс 403/348)
Поворотный обвязочный кулачок или клин (Класс 403/350)
Собирается множественными манипуляциями с членом (класс 403/353)
Диаметральный концевой паз является соединительным элементом (Класс 403/354)
Включая отдельно вставляемый ключ (Класс 403/355)
- Стержень с продольными шлицами или канавками (Класс 403/359. 1)
Стержень с внешним заплечиком или головкой (Класс 403/360)
Розетка (класс 403/361)
Включая установочный винт (Класс 403/362)
Промежуточные уголки или швеллеры (Класс 403/363)
Взаимноконечные концы (Класс 403/364)
Прокладка или втулка с радиальной вставкой (Класс 403/365)
Зажимные элементы (Класс 403/373)
Периферийное расширение, углубление или прорезь на одном элементе являются компонентом сустава (класс 403/375).
Включая отдельный фиксатор (класс 403/376)
Увеличенная головка в дополнительном углублении, например, ласточкин хвост и т. д. (класс 403/381)
Жесткий угол (Класс 403/382)
Некруглое сечение стержня является соединительным элементом (класс 403/383)
Сторона стержня к пластине или стороне (Класс 403/384)
Соединение двойным зажимом (Класс 403/385)
Скоба является неотъемлемой частью или независимо прикреплена к ближней стороне пластины или другой стороне (класс 403/386)
Фланцевый или рифленый стержень (Класс 403/387)
Проходит через соединитель (Класс 403/388)
Стержни с боковым расположением (класс 403/389)
Соединены гибкой стяжкой (Класс 403/392)
Притертые концы стержней (Класс 403/393)
Диагональный разъем (Класс 403/394)
Один стержень, удерживаемый между выступом, и другой стержень, проходящий через отверстие в ножке соединителя (класс 403/395)
Два стержня с одним соединителем (класс 403/39).
6)Эластичный зажим (Класс 403/397)
Хомут или соединитель кольцевого типа (класс 403/398)
Скрещенные стержни (Класс 403/400)
Соединение под углом (Класс 403/401)
Соединительные перемычки внутренней или внешней периферии (класс 403/402)
Жесткий уголок (Класс 403/403)
Особая конфигурация или материал элемента (класс 403/404)
Соединительный элемент, проходящий через совмещенные отверстия во множественных элементах (класс 403/408. 1)
Имеющие кулачок, клин или коническую часть (Класс 403/409.1)
Разное (класс 403/410)
Коллекции иностранных патентов
Иностранные документы, относящиеся к классу (Класс 403/FOR000)
Шарнирные члены
Блокируется в фиксированном положении
Телескопические элементы (403/109) (Класс 403/FOR100)
Наличие рабочего механизма (403/322) (Класс 403/FOR101)
Интерфитированные члены
Включая дискретный фиксатор
Дайджесты
Магнитный (Класс 403/DIG1)
Обработка металла (Класс 403/DIG2)
Расцепитель, реагирующий на нагрузку (класс 403/DIG3)
Быстросъемный (Класс 403/DIG4)
Угольный электрод (Класс 403/DIG5)
Возвратно-поступательный шариковый фиксатор (Класс 403/DIG6)
Разрезное кольцо или абатмент (класс 403/DIG7)
Радиальный кулачок или эксцентрик (Класс 403/DIG8)
Регулируемый зажим (Класс 403/DIG9)
Легко разъединяемые стыки панелей (Класс 403/DIG10)
Тип мебели с защелкой (Класс 403/DIG11)
Тип мебели, имеющий вращающийся крепежный элемент или крепежный элемент, затягивающий соединение (Класс 403/DIG12)
Тип мебели (Класс 403/DIG13)
Эластичная защелка, защелка или фиксатор (класс 403/DIG14)
Соединительные пластины для коллинеарных стержней (Класс 403/DIG15)
Epac2 в дофаминовых нейронах среднего мозга способствует подкреплению кокаина за счет усиления высвобождения дофамина
Abstract
Повторяющееся воздействие наркотиков, вызывающих зависимость, приводит к усилению передачи сигналов цАМФ в мезолимбической дофаминовой системе, молекулярная адаптация, которая, как считается, играет решающую роль в развитии лекарственной зависимости. Обменный белок, непосредственно активируемый цАМФ (Epac2), является основным эффектором цАМФ, обильно экспрессируемым в головном мозге. Однако остается неизвестным, способствует ли Epac2 усилению кокаина. Здесь мы сообщаем, что Epac2 в мезолимбической дофаминовой системе способствует усилению приема кокаина за счет усиления высвобождения дофамина. Условный нокаут Epac2 из дофаминовых нейронов среднего мозга (Epac2-cKO) и селективный ингибитор Epac2 ESI-05 снижали самостоятельный прием кокаина у мышей как при фиксированном, так и при прогрессивном соотношении подкрепления, а также в широком диапазоне доз кокаина. Кроме того, Epac2-cKO приводил к уменьшению вызванного высвобождения дофамина, тогда как агонизм Epac2 резко усиливал высвобождение дофамина в прилежащем ядре in vitro. Этот механизм играет центральную роль в поведенческих эффектах нарушения Epac2, поскольку хемогенетическая стимуляция дофаминовых нейронов вентральной области покрышки (VTA) посредством индуцированной дезхлорклозапином (DCZ) активации Gs-DREADD увеличивает высвобождение дофамина и обращает вспять нарушение самостоятельного приема кокаина в Epac2. -cKO мыши. И наоборот, хемогенетическое ингибирование дофаминовых нейронов VTA с помощью Gi-DREADD уменьшало высвобождение дофамина и самостоятельный прием кокаина у мышей дикого типа. Таким образом, Epac2-опосредованное усиление высвобождения дофамина может представлять собой новый и мощный механизм, который способствует кокаиновому подкреплению.
Оценка редактора
В этой рукописи сообщается, что Epac2, нижестоящий эффектор цАМФ, позитивно регулирует кокаиновое вознаграждение, изменяя свойства высвобождения дофамина в стриатуме. В исследовании используется широкий спектр технических подходов для тщательной характеристики клеточных и поведенческих ролей Epac2 у мышей. Вместе эти результаты обеспечивают убедительное важное понимание Epac2 как потенциальной пресинаптической молекулярной мишени, с помощью которой можно манипулировать передачей сигналов дофамина и приемом лекарств, и представляет интерес для ученых, изучающих передачу дофамина и расстройства, связанные с употреблением психоактивных веществ.
https://doi.org/10.7554/eLife.80747.sa0
- Письмо-решение
- Обзоры на науку
- Процесс проверки eLife
Введение
Вознаграждающие эффекты кокаина в первую очередь опосредованы усиленной дофаминергической передачей в мезолимбической дофаминовой системе (Anderson and Pierce, 2005). Дофамин D 1 — и D 2 -подобные рецепторы связаны с белками G αs и G αi/o , что приводит к увеличению или уменьшению внутриклеточного цАМФ, соответственно (Tritsch and Sabatini, 2012). Повторяющееся воздействие кокаина вызывает активацию передачи сигналов цАМФ в вентральной области покрышки (VTA) и прилежащем ядре (NAc) (Nestler, 2001; Self, 2004; Nestler, 2016). Было показано, что в ВТА повторное воздействие кокаина снижает экспрессию G αi/o (Nestler et al. , 1990; Striplin and Kalivas, 19).92), что приводит к повышению уровня цАМФ. Протеинкиназа A (PKA) и обменный белок, непосредственно активируемый цАМФ (Epac), являются двумя основными внутриклеточными эффекторами цАМФ (de Rooij et al., 1998; Kawasaki et al., 1998; Tritsch and Sabatini, 2012) и имеют многочисленные нижележащие сигнальные мишени. , тем самым преобразовывая изменения уровней цАМФ в клеточные адаптации, которые могут способствовать поведенческим изменениям. Сигнальный путь цАМФ-PKA-CREB (белок, связывающий элемент ответа цАМФ) широко изучался в контексте наркомании (Nestler, 2001; Self, 2004; Anderson and Pierce, 2005; Nestler, 2016). Тем не менее, в нескольких исследованиях изучалось, как «другой» эффектор цАМФ, Epac, регулирует клеточные и поведенческие эффекты наркотиков, вызывающих зависимость.
Существуют две изоформы Epac, Epac1 и Epac2. Epac2 преимущественно экспрессируется в головном мозге, тогда как экспрессия Epac1 в головном мозге очень низка (de Rooij et al. , 1998; Kawasaki et al., 1998). Делая записи целых клеток на срезах ex vivo, полученных от мышей с глобальным нокаутом Epac1 и Epac2 (Yang et al., 2012), мы показали, что Epac2, но не Epac1, регулирует адаптацию возбуждающей и тормозной синаптической пластичности в VTA-нейронах, индуцированных дофамином. непредвиденным воздействием кокаина (Liu et al., 2016; Tong et al., 2017). Самостоятельный прием лекарств обеспечивает меру подкрепляющей эффективности наркотиков, вызывающих злоупотребление, и мотивацию животного к получению полезных наркотиков (O’Connor et al., 2011; Hiranita, 2015). Общая цель этого исследования заключалась в том, чтобы определить, регулирует ли передача сигналов Epac2 в дофаминовых нейронах среднего мозга самостоятельный прием кокаина. Активаторы Epac усиливают пресинаптическое высвобождение глутамата за счет увеличения ассоциации между белками активной зоны Rab3A, RIM и Munc13, тем самым облегчая стыковку везикул и экзоцитоз (Ozaki et al., 2000; Ferrero et al., 2013; Fernandes et al. , 2015). Аксональное высвобождение дофамина в стриатуме опосредовано везикулярным экзоцитозом и требует наличия активных зоноподобных механизмов (Liu and Kaeser, 2019).). Специфический для дофаминовых нейронов нокаут RIM отменяет вызванное потенциалом действия высвобождение дофамина в стриатуме (Liu et al., 2018a). Однако ни в одном из предыдущих исследований не изучалось, модулирует ли Epac2 высвобождение дофамина.
В настоящем исследовании мы создали мышей с условным нокаутом Epac2 (Epac2-cKO), специфичных для дофаминовых нейронов, и использовали Epac2-селективный антагонист ESI-05 (Tsalkova et al., 2012) для изучения роли передачи сигналов Epac2 в дофаминерге среднего мозга. нейроны в регуляции кокаинового подкрепления. Мы изучили влияние ESI-05 и Epac2-cKO на схемы подкрепления с фиксированным соотношением-1 (FR1) и прогрессивным соотношением (PR) и кривые доза-реакция кокаина. Поскольку подкрепляющая эффективность кокаина в первую очередь опосредована его эффектом повышения уровня внеклеточного дофамина (Anderson and Pierce, 2005), мы проверили гипотезу о том, что Epac2 может способствовать подкреплению кокаином путем модуляции высвобождения дофамина. Сначала мы определили, как фармакологические и генетические манипуляции с Epac2 влияют на высвобождение дофамина в полосатом теле. Затем мы экспрессировали Gs- и Gi-дизайнерные рецепторы, активируемые исключительно дизайнерскими наркотиками (DREADD) (Roth, 2016) в дофаминовых нейронах VTA, и исследовали, влияет ли двунаправленная модуляция высвобождения дофамина с активацией DREADD на самостоятельный прием кокаина у дикого типа (WT). ) и мышей Epac2-cKO. Вместе наши исследования предоставляют доказательства критической роли Epac2 в регуляции высвобождения дофамина и кокаинового подкрепления.
Результаты
Ингибирование Epac2 снижает самостоятельный прием кокаина при множественных схемах подкрепления и дозах кокаина
Мы исследовали влияние селективного ингибитора Epac2 ESI-05 на самостоятельный прием кокаина у примерно равного количества самцов и самок мышей с использованием системы самостоятельного введения Instech, которая значительно продлевает срок службы катетера (Slosky et al. , 2020; Слоски и др., 2021). Мышей C57BL/6J обучали самостоятельному введению кокаина по схеме подкрепления FR1 (0,5 мг/кг/инфузия) в течение 3-часовых сеансов в течение 10 дней, и 18 из 20 мышей с незаращенными катетерами приобрели стабильное самостоятельное введение кокаина (критерии подробно описаны). в материалах и методах). Среднее количество сеансов для достижения стабильного приобретения составило 5,4 ± 0,3 сеанса (рис. 1А). В последующие дни мышам вводили ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) или носитель (0 мг/кг, внутрибрюшинно) за 10 минут до тестирования на самостоятельное введение в дизайне Latin Square. ESI-05 дозозависимо уменьшал активные тычки в нос (рис. 1B) и инфузии кокаина (рис. 1C), но не оказывал значительного влияния на неактивные тычки в нос (рис. 1D; подробная статистика для этого и последующих результатов описана в подписях к рисункам). Мыши, получавшие носитель, демонстрировали неизменно высокую скорость вливаний кокаина с преимущественно регулярными интервалами в течение всего 3-часового сеанса; ESI-05 дозозависимо снижал скорость введения кокаина, которая характеризовалась неравномерными интервалами и спорадическим приемом, особенно при более высокой дозе ESI-05 (10 мг/кг, рисунок 1 — приложение к рисунку 1). Как и в предыдущем исследовании (Slosky et al., 2021), никаких существенных различий по полу не наблюдалось ни по одному показателю, поэтому в последующих исследованиях данные были объединены по половому признаку.
ESI-05 дозозависимо снижал самостоятельный прием кокаина при схемах подкрепления с фиксированным соотношением-1 (FR1) и прогрессивным соотношением (PR) у мышей.
( A ) Активные уколы в нос и неактивные уколы в нос и инфузии кокаина во время 10-дневного обучения самостоятельному введению FR1 (0,5 мг/кг/инфузия, n = 18, 9 мужчин и 9 женщин). График FR1 имеет короткий период ожидания (10 с) после каждого активного тыкания в нос, в течение которого дополнительные тычки в нос не приводят к вливанию кокаина. В результате количество уколов носом превышает количество вливаний кокаина. ( B–D ) Среднее количество активных уколов носом (однофакторный дисперсионный анализ: F 2,32 = 30,9, p<0,001), инъекций кокаина (однофакторный дисперсионный анализ: F 2,32 =31,9, p<0,001) и неактивные тычки в нос (однофакторный ANOVA: F 2,32 = 0,5, p=0,613) в ответ на предварительную обработку ESI-05 (0, 3,3, 10 мг/кг , внутрибрюшинно) по схеме подкрепления FR1 (n=11–13, 6 мужчин и 5–7 женщин в группе). ( E–F ) Предварительное лечение ESI-05 привело к значительному сдвигу вниз на кривой доза-реакция для инфузий кокаина (двухфакторный дисперсионный анализ с повторными измерениями: лечение ESI-05, Ж 2,22 = 45,5, р<0,001; доза кокаина, F 6132 = 51,0, p<0,001; Лечение ESI-05 × взаимодействие дозы кокаина, F 12 132 = 6,5, p<0,001; 3,3 против 0 мг/кг ESI-05, красный *; 10 против 0 мг/кг ESI-05, фиолетовый *; 3,3 по сравнению с 10 мг/кг ESI-05 черный # ) и снижение общего потребления кокаина (двухфакторный дисперсионный анализ с повторными измерениями: лечение ESI-05, F 2,22 = 66,5, p<0,001; кокаин доза, F 6132 = 498,1, р<0,001; ESI-05 × взаимодействие дозы кокаина, F 12 132 = 6,7, p<0,001; 3,3 против 0 мг/кг ESI-05, красный *; 10 против 0 мг/кг ESI-05 фиолетовый *; 3,3 против 10 мг/кг ESI-05 black # ) на кривой доза-реакция (n=8–9, 5 мужчин и 4 женщины). ( G–H ) ESI-05 дозозависимо снижал количество инъекций кокаина (односторонний ANOVA: F 2,28 = 16,1, p<0,001; n=10–11, 5 мужчин и 5 –6 женщин) и точка останова (равная дисперсия Брауна-Форсайта: p<0,05; однофакторный дисперсионный анализ Краскела-Уоллиса по рангам: ESI-05, Н =17,5, р<0,001; n=10–11, 5 мужчин и 5–6 женщин) по схеме PR-подкрепления. *р<0,05, **р<0,01, ***р<0,001; # p<0,05, ## p<0,01, ### p<0,001 (см. также рис. 1 — приложение 1 к рисунку).
Затем мы исследовали влияние ESI-05 на самостоятельный прием всего диапазона доз кокаина у мышей, которые научились самостоятельно принимать кокаин в течение 10 дней после 10-дневного обучения по схеме FR1. При использовании начальной 0,5 мг/кг/инфузия с последующим снижением доз за один сеанс (1,0, 0,25, 0,125, 0,0625, 0,03125 и 0,0 мг/кг/инфузия; см. Материалы и методы) (Anderson et al., 2018) , мы наблюдали классические перевернутые U-образные кривые доза-реакция у мышей C57BL/6J, при этом максимальная инфузия происходила при дозе 0,125 мг/кг/инфузия (рис. 1E). После того, как мыши достигли стабильного поведения при самостоятельном введении в этой парадигме многократного введения, за 10 минут до самостоятельного введения кокаина вводили ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) или носитель (0 мг/кг). Предварительная обработка ESI-05 привела к дозозависимым сдвигам кривой доза-реакция кокаина (рис. 1E) и ослаблению потребления кокаина (рис. 1F).
Затем мы определили, влияет ли ESI-05 на мотивацию к самостоятельному приему кокаина в соответствии с графиком PR-подкрепления (Richardson and Roberts, 1996). Мыши сначала подвергались самостоятельному введению кокаина в течение 10 дней по схеме FR1, как описано выше, а затем были переведены на схему PR (0,5 мг/кг/инфузия). Когда пограничная точка PR стабилизировалась, изучали влияние ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) или носителя на инфузии кокаина и пограничную точку. Предварительная обработка ESI-05 дозозависимо снижала инфузии кокаина (рис. 1G) и контрольную точку (рис. 1H) по сравнению с носителем. Таким образом, ESI-05 снижал мотивацию к получению кокаина, когда требуемое усилие постепенно увеличивалось. В совокупности приведенные выше результаты показывают, что ESI-05 снижает потребление кокаина и/или мотивацию к употреблению кокаина.
Специфический нокаут дофаминовых нейронов нарушенного Epac2 Самостоятельный прием кокаина
Затем мы определили, влияет ли условное удаление Epac2 из дофаминовых нейронов среднего мозга на самостоятельный прием кокаина. Мышей с нокаутом Epac2 по дофаминовым нейронам (Epac2-cKO) получали путем скрещивания гомозиготных мышей Epac2 loxP/loxP с гетерозиготными мышами Slc6a3 IRES cre/+ . Мыши Epac2-cKO ( Slc6a3 IRES cre/+ /Epac2 loxP/loxP ) and WT controls ( Slc6a3 IRES cre/+ /Epac2 wt/wt or Slc6a3 +/+ /Epac2 loxP/loxP ) были в целом нормальными с точки зрения выживаемости, внешнего вида, массы тела (рис. 2 — дополнение к рисунку 1A) и исходной двигательной активности (рис. 2 — дополнение к рисунку 1B). Мы подтвердили селективный нокаут Epac2 из дофаминовых нейронов VTA с помощью одноклеточной обратной транскриптазы-ПЦР (scRT-PCR). Записи возбуждения потенциала действия, прикрепленные к клеткам, были сделаны в нейронах VTA с последующей записью целых клеток и сбором содержимого цитоплазмы для scRT-PCR. Профили экспрессии генов нейронов характеризовались обнаружением мРНК для Epac1 , Epac2 , тирозингидроксилаза ( Th , маркер дофаминовых нейронов), глутаматдекарбоксилаза 2 ( Gad2 , маркер нейронов ГАМК), семейство переносчиков растворенного вещества 17 (везикулярный переносчик глутамата), член 6 (4 Slc, 4 Slc глутаматный нейронный маркер) и ген домашнего хозяйства глицеральдегид-3-фосфатдегидрогеназы ( Gapdh ). Мы обнаружили, что Epac2 экспрессировался как в Th + , так и в Gad2 + 9. 1352 нейрона в VTA мышей WT (рис. 2 — дополнение к рисунку 2A), но было обнаружено только в нейронах Gad2 + , а не в нейронах Th + в VTA мышей Epac2-cKO (рис. 2— дополнение к рисунку 2В). Epac1 редко экспрессировался в нейронах мышей WT и Epac2-cKO, что согласуется с низкой экспрессией Epac1 в головном мозге (Kawasaki et al., 1998). Мы также сравнили электрофизиологические свойства дофаминовых нейронов VTA от мышей WT и Epac2-cKO и не обнаружили существенных различий в частоте срабатывания потенциала действия или емкости мембраны (рис. 2 — дополнение к рисунку 3).
Затем мы исследовали, изменился ли самостоятельный прием кокаина у мышей Epac2-cKO. Мышей Epac2-cKO и контрольную группу дикого типа обучали самостоятельному введению кокаина по графику FR1, как описано выше. Не было существенной разницы между процентом мышей в каждом генотипе, которые приобрели стабильное самостоятельное введение кокаина после 10 сеансов, с 90,9% (10 из 11) мышей WT и 84,6% (11 из 13) мышей Epac2-cKO с открытыми катетерами. отвечающие критериям стабильного самоуправления (χ 2 =0,024, р=0,877). Из мышей, которые не соответствовали критериям, одна мышь WT и одна мышь Epac2-cKO продемонстрировали <20 вливаний кокаина, в то время как другая мышь Epac2-cKO показала большое количество вливаний кокаина (> 50), но не смогла развить активный ответ по сравнению с неактивным. дискриминация. У мышей, которые научились стабильному самоуправлению, среднее количество сеансов обучения для достижения критериев существенно не отличалось между генотипами (WT, 5,4 ± 0,5 сеанса; Epac2-cKO, 6,8 ± 0,7 сеанса; 9).0004 t 19 =1,6, р=0,130). Тем не менее, у мышей Epac2-cKO наблюдалось значительное снижение количества активных движений носом (рис. 2А) и инъекций кокаина (рис. 2В). Мыши WT, как правило, демонстрировали высокую скорость реакции с преимущественно регулярными интервалами, в то время как мыши Epac2-cKO демонстрировали паттерн, напоминающий паттерн мышей, получавших ESI-05, характеризующийся спорадическим потреблением с вкраплениями длительных промежутков (рис. 2 — дополнение к рисунку 4). Неактивные тычки в нос существенно не различались между генотипами (рис. 2С). Таким образом, хотя мыши WT и Epac2-cKO сходным образом научились самостоятельно принимать стабильный кокаин, у мышей Epac2-cKO наблюдалось значительно сниженное потребление кокаина.
Специфический нокаут дофаминовых нейронов Epac2 нарушил усвоение кокаина при самостоятельном введении и усилил эффективность.
( A ) Мыши с условным нокаутом Epac2 (Epac2-cKO) демонстрировали значительно более низкие активные тычки носом во время обучения самостоятельному введению с фиксированным отношением 1 (FR1) (двухфакторный дисперсионный анализ с повторными измерениями: генотип, F 1,19 = 8,2, р=0,010; тренировка, Ж 9,171 = 10,0, р<0,001; генотип × взаимодействие тренировочных занятий, F 9,171 = 0,7, p = 0,670; n=10–11, по 5 мужчин и 5–6 женщин в группе). ( B ) Количество инъекций кокаина во время тренировки было значительно снижено с помощью Epac2-cKO (двусторонний дисперсионный анализ с повторными измерениями: генотип, F 1,19 = 8,3, p = 0,010; тренировочная сессия, F 9171 = 13,3, p<0,001, генотип × взаимодействие тренировочных занятий, Ж 9171 = 0,9, р=0,543; п=10–11). ( C ) Не было существенной разницы в неактивных тычках носом между генотипами (двусторонний дисперсионный анализ с повторными измерениями: генотип, F 1,19 = 1,4, p = 0,247; n = 10–11, 5 самцов и 5-6 женщин в группе). ( D – E ) У мышей Epac2-cKO наблюдался сдвиг кривой доза-реакция кокаина вниз (двусторонний дисперсионный анализ с повторными измерениями: генотип, F 1,15 = 20,0, p <0,001; доза кокаина , Ж 6,90 = 49,4, р<0,001; взаимодействие генотип × доза кокаина, F 6,90 = 2,2, р=0,046; n=8–9, 4–5 мужчин и 4 женщины в группе) и снижение потребления кокаина (двусторонний дисперсионный анализ с повторными измерениями: генотип, F 1,15 = 10,4, p = 0,006; доза кокаина, F 6,90 = 62,0, p<0,001, взаимодействие генотипа и дозы кокаина, F 6,90 = 2,5, p=0,025, n=8–9, 4 мужчины и 4–5 женщин в группе ). ( F–G ) Мыши Epac2-cKO продемонстрировали меньшее количество инъекций кокаина ( t -тест: t 19 = 2,6, p = 0,019) и более низкую точку разрыва ( t -тест: t 19 = 2,7, p = 0,015) по схеме подкрепления с прогрессивным соотношением (PR) (n = 9–11, 5 мужчин и 4–6 женщин в группе). *p<0,05, **p<0,01, ***p<0,001 (см. также рис. 2 — приложения к рисунку 1–4).
Чтобы проверить гипотезу о том, что Epac2-cKO подавляет подкрепляющие эффекты кокаина, мы исследовали самостоятельный прием кокаина в парадигме множественных доз с использованием той же когорты мышей WT и Epac2-cKO, как показано на рис. 2A–C. У мышей Epac2-cKO наблюдался значительный сдвиг кривой доза-реакция кокаина по сравнению с мышами дикого типа (рис. 2D). Мыши Epac2-cKO также потребляли меньше кокаина, чем мыши WT (рис. 2E). Наконец, мы исследовали, изменились ли у мышей Epac2-cKO побудительные мотивы для получения вознаграждения кокаином в соответствии со схемой подкрепления PR в отдельной когорте мышей. Мышей WT и Epac2-cKO подвергали самостоятельному введению кокаина FR1 в течение 10 дней, а затем переводили на схему PR, как описано выше. Мыши Epac2-cKO получали значительно меньше инфузий кокаина (рис. 2F) и имели значительно сниженную точку разрыва (рис. 2G). В совокупности эти результаты свидетельствуют о том, что у мышей Epac2-cKO снижается мотивация приобретать кокаин при многократном приеме кокаина и при постепенном увеличении требуемых усилий.
Ингибирование Epac2 или Epac2-cKO не оказывает значительного влияния на самостоятельный пероральный прием сахарозы
Способствует ли активность Epac2 специфическому подкреплению наркотиками или ее можно обобщить на другие поощрения, не связанные с наркотиками? Чтобы проверить это, мы исследовали влияние системного ингибирования Epac2 и условного нокаута в дофаминовых нейронах на самостоятельный прием перорального вознаграждения сахарозой как по схемам FR1, так и по схемам PR. Мышей обучали самостоятельному пероральному введению сахарозы по схеме FR1 (10% сахарозы, 35 мкл) в течение 1 часа в течение 10 дней (рис. 3 — дополнение к рисунку 1A). В последующие дни мышам вводили ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) или носитель за 10 минут до тестирования самостоятельного введения в дизайне Latin Square. ESI-05 не оказывал значительного влияния на активные и неактивные тычки в нос или потребление сахарозы (рис. 3 — дополнение к рисунку 1B-D). Отдельную группу мышей обучали самостоятельному введению сахарозы в соответствии с графиком подкрепления PR. Опять же, ESI-05 не влиял на потребление сахарозы или контрольную точку (Рисунок 3 — дополнение к рисунку 1E-F) при этой схеме подкрепления.
Затем мы исследовали, изменяет ли Epac2-cKO самостоятельный пероральный прием сахарозы при схемах подкрепления FR1 и PR. При схеме FR1 не было значительного влияния генотипа на активные и неактивные тычки в нос или потребление сахарозы в течение 10-дневного сеанса (рис. 3А–С). Как WT, так и мыши Epac2-cKO демонстрировали преимущественно регулярное потребление сахарозы в течение всего 1-часового сеанса, хотя всплески высоких скоростей потребления сахарозы также можно было обнаружить у обоих генотипов (рис. 3 — дополнение к рисунку 2). Точно так же Epac2-cKO не влиял на потребление сахарозы или контрольную точку при схеме PR (рис. 3D-E). Взятые вместе, эти результаты показывают, что ингибирование и делеция Epac2 существенно не изменили подкрепляющие эффекты приятного немедикаментозного вознаграждения.
Условный нокаут Epac2 (Epac2-cKO) не влиял на самостоятельный прием сахарозы при схемах подкрепления с фиксированным соотношением-1 (FR1) и прогрессивным соотношением (PR) у мышей.
( A – C ) Epac2-cKO не влиял на усвоение сахарозы при самостоятельном введении после 10 дней обучения по графику FR1 (двусторонний анализ с повторными измерениями: тренировочная сессия по активным тычкам в нос, Ж 9,126 = 20,8, р<0,001; тренировка по неактивным тычкам в нос, Ж 9,126 = 6,4, р<0,001; тренировка по приему сахарозы, Ж 9,126 = 21,3, р<0,001; генотип по активным тычкам носом, F 1,14 = 0,07, р=0,802; генотип по неактивным тычкам носа, F 1,14 = 3,1, р=0,099; генотип по сахарозному вознаграждению, F 1,14 = 0,08, р=0,787; тренировка × взаимодействие генотипов при активных тычках носом, Ж 9126 = 0,5, р=0,849; при неактивных тычках в нос F 9,126 = 1,5, р=0,151; потребление сахарозы, Ж 9,126 = 0,6, р=0,8). ( D–E ) Epac2-cKO не влиял на среднее количество потребления сахарозы ( t -test: t 14 =0,4, p=0,730) или контрольные точки (ранг Манна-Уитни: U =0,6, р=0,561) по схеме PR. n=8, по 4 мужчины и 4 женщины на группу (см. также рис. 3 — приложение 1 к рисунку).
Epac2-cKO приводил к снижению высвобождения дофамина в стриатуме in vitro
Какими могут быть механизмы, с помощью которых Epac2 в дофаминовых нейронах регулирует самостоятельный прием кокаина? Злоупотребление наркотиками, включая кокаин, повышает уровень дофамина в NAc, что, как считается, лежит в основе их подкрепляющего действия (Di Chiara and Imperato, 1988; Koob and Bloom, 1988). Epac2 взаимодействует с белками активной зоны RIM и Munc13, чтобы облегчить экзоцитоз глутамата из возбуждающих синапсов (Ozaki et al., 2000; Tengholm and Gylfe, 2017). Условный нокаут RIM из дофаминовых нейронов среднего мозга отменял высвобождение дофамина, запускаемое потенциалом действия (Liu et al. , 2018a). Одна возможность заключается в том, что нарушение Epac2 ослабляет высвобождение дофамина из окончаний аксонов. Чтобы проверить это, мы использовали циклическую вольтамперометрию с быстрым сканированием (FSCV) для мониторинга вызванного электричеством высвобождения дофамина в срезах полосатого тела у мышей WT и Epac2-cKO. Записи FSCV были сделаны в ядре NAc (сайт 1) и оболочке (сайт 2), дорсомедиальном стриатуме (DMS, сайт 3) и дорсолатеральном полосатом теле (DLS, сайт 4; рисунок 4A и D). Дорсальное полосатое тело преимущественно иннервируется дофаминовыми нейронами в компактной части черной субстанции (SNc), в то время как NAc преимущественно иннервируется дофаминовыми нейронами VTA (Ogawa and Watabe Uchida, 2018). Фиксированная интенсивность стимуляции (250 мкА, продолжительность 0,2 мс) либо с одним электрическим стимулом, либо с серией стимулов (5 импульсов, 100 Гц) применялась для стимуляции высвобождения дофамина. Пиковые амплитуды вызванного выброса дофамина в ответ на одиночный импульс и высокочастотную импульсную стимуляцию были значительно снижены во всех субрегионах полосатого тела, взятых в срезах мышей Epac2-cKO, по сравнению с мышами WT (рис. 4A, B, D и E). Обратный захват дофамина транспортером дофамина (DAT) является основным механизмом, ответственным за внеклеточный клиренс дофамина в головном мозге (Gainetdinov et al., 19).98; Бенуа-Маранд и др., 2000). Константа экспоненциального затухания τ вызванных ответов на дофамин обеспечивает надежную меру для мониторинга изменений обратного захвата дофамина (Yorgason et al., 2011). Мы обнаружили, что не было значительных изменений τ ни от одного импульса, ни от высокочастотной импульсной стимуляции во всех субрегионах полосатого тела, отобранных в срезах мышей Epac2-cKO, по сравнению с мышами WT (рис. 4A, C, D и F). Таким образом, у мышей Epac2-cKO было снижено вызванное высвобождение дофамина без значительного изменения обратного захвата дофамина.
Нокаут
Epac2 снижал вызванное высвобождение дофамина в стриатуме in vitro.
( A ) Среднее значение концентрации дофамина в зависимости от времени в четырех различных субрегионах полосатого тела (ядро прилежащего ядра [NAc], оболочка NAc, дорсально-медиальное полосатое тело [DMS] и дорсально-латеральное полосатое тело [DLS]), вызванное одним электрическим импульсом. ( B ) По сравнению с мышами дикого типа (WT) у мышей с условным нокаутом Epac2 (Epac2-cKO) наблюдалось снижение вызванного выброса дофамина во всех субрегионах полосатого тела в ответ на одиночную импульсную стимуляцию ( t -испытания: керн NAc, t 36 =2,3, р=0,028; оболочка NAc, т 36 =4,4, р<0,001; ДМС, т 41 =7,4, р<0,001; ДЛС, т 39 =6,1, р<0,001; n=18–22 среза от 4 до 5 самцов и самок мышей). ( C ) Не было выявлено существенных различий в константах времени затухания между мышами WT и Epac2-cKO при одноимпульсной стимуляции во всех участках регистрации ( t -тесты: ядро NAc, т 36 =0,9, р=0,366; оболочка NAc, т 36 =0,2, р=0,842; ДМС, т 41 =0,3, р=0,736; ДЛС, т 39 =0,4, р=0,676; n=18–22 среза от 4 до 5 самцов и самок мышей). ( D ) Среднее значение концентрации дофамина в зависимости от времени в четырех различных субрегионах полосатого тела в ответ на 5-импульсную электрическую стимуляцию с частотой 100 Гц. ( E ) По сравнению с мышами WT, у мышей Epac2-cKO наблюдалось снижение вызванного выброса дофамина во всех субрегионах полосатого тела в ответ на 5-импульсную электрическую стимуляцию ( t -испытания: керн NAc, t 29 =2,4, р=0,025; оболочка NAc, т 28 =3,4, р=0,002; ДМС, т 31 =6,3, р<0,001; ДЛС, т 29 =5,3, р<0,001; n=14–18 срезов от 4 до 5 самцов и самок мышей). ( F ) Не было выявлено существенных различий в константах времени затухания между мышами WT и Epac2-cKO при 5-импульсной электростимуляции ( t — тесты: ядро NAc, t 29 = 0,006, р = 0,999; оболочка NAc, т 28 =0,3, р=0,770; ДМС, т 31 =0,3, р=0,801; ДЛС, т 29 =0,1, р=0,918; n=14–18 срезов от 4 до 5 самцов и самок мышей). *р<0,05, **р<0,01, ***р<0,001.
Активация Epac и PKA независимо увеличивала высвобождение дофамина в NAc in vitro
Чтобы непосредственно проверить влияние активации Epac2 на высвобождение дофамина в оболочке NAc, мы отслеживали высвобождение дофамина, вызванное одиночным электрическим стимулом, который применялся с 2-минутными интервалами. Sp-8-BnT-cAMPS («S-220») представляет собой Epac2-селективный агонист со специфичностью в отношении Epac2 по сравнению с PKA и Epac1 (Schwede et al., 2015). Применение ванны с S-220 приводило к зависимому от концентрации увеличению вызванного высвобождения дофамина в срезах WT (рис. 5A-B), которое блокировалось Epac2-cKO (рис. 5C) и ESI-05 (20 мкМ, рис. 5D). Напротив, ингибитор ПКА H89(1 мкМ) не имел никакого эффекта (рис. 5Е). Наконец, мы исследовали, изменяет ли активация PKA высвобождение дофамина в NAc. 6-Bnz-cAMP является мембранопроницаемым и селективным активатором PKA и не активирует Epac (Christensen et al., 2003; Hewer et al., 2011). Применение ванны с 6-Bnz-цАМФ (1 мкМ) увеличивало высвобождение дофамина в оболочке NAc в срезах мышей дикого типа (рис. 5F). Этот эффект блокировался H89, но не изменялся Epac-cKO (фиг. 5F). Таким образом, активация путей Epac2 и PKA независимо друг от друга увеличивает вызванное высвобождение дофамина.
Активация Epac2 увеличивала высвобождение дофамина в прилежащем ядре (NAc) in vitro.
( A ) Высвобождение дофамина было заметно увеличено при перфузии S-220 (10 мкМ) в срезах NAc от мышей дикого типа (WT) (спаренные t -test; t 8 = 9,1 , p<0,001; n=9 срезов от 3 до 4 самцов и самок мышей). ( B ) S-220 увеличивал высвобождение дофамина дозозависимым образом в срезах NAc от мышей дикого типа (односторонний ANOVA: F 3,27 = 20,4, p<0,001, n=6–9 срезов из от 3 до 4 самцов и самок мышей). ( C ) S-220 (10 мкМ) не увеличивал высвобождение дофамина в срезах NAc от мышей с условным нокаутом Epac2 (Epac2-cKO) (спаренные t — тест: t 9 = 0,4, p = 0,680 , p=0,236; n=9 срезов от 3 до 4 самцов и самок мышей). ( D ) Предварительная инкубация срезов с ESI-05 (20 мкМ) блокировала S-220-индуцированное увеличение высвобождения дофамина ( t -тест: t 16 =7,4, p<0,001; n=9 срезов от 3 до 4 самцов и самок мышей). ( E ) Предварительная инкубация срезов с H89 (1 мкМ) не изменила индуцированное S-220 увеличение высвобождения дофамина ( t — тест: t 15 = 1,47, p = 0,162; n = 8 –9 срезов от 3 до 4 самцов и самок мышей). ( F ) 6-Bnz (1 мкМ) увеличивает высвобождение дофамина в срезах NAc, полученных от мышей дикого типа (спаренные t — тест: t 8 =6,5, р<0,001; n=9 срезов от 3 до 4 самцов и самок мышей). Увеличение блокировалось H89 (1 мкМ; t -тест: t 14 = 5,1, p<0,001; n=7–9 срезов от 3 до 4 самцов и самок мышей). 6-Bnz также индуцировал увеличение высвобождения дофамина в срезах мышей Epac2-cKO (пара t -test: t 8 = 6,5, p<0,001; n = 8 срезов от 3 до 4 самцов и самок мышей). ), который достоверно не отличался от мышей дикого типа ( t -test: т 15 =0,9, р=0,408; n=8–9 срезов от 3 до 4 самцов и самок мышей). ***р<0,001.
Хемогенетическое усиление высвобождения дофамина увеличивает самостоятельный прием кокаина у мышей Epac2-cKO
Если снижение высвобождения дофамина в результате разрушения Epac2 способствует нарушению самостоятельного приема кокаина, избирательное усиление высвобождения дофамина может обратить вспять эти поведенческие эффекты. Мы проверили, может ли хемогенетическое усиление высвобождения дофамина из дофаминовых нейронов VTA восстановить самостоятельный прием кокаина у мышей Epac2-cKO. AAV8-hSyn-DIO-rM3D(Gs)-mCherry или AAV8-hSyn-DIO-mCherry билатерально микроинъецировали в VTA Epac2-cKO ( Slc6a3 IRES cre/+ / Epac2 loxP/loxP ) мышей. Через 2–3 недели экспрессия трансгена была подтверждена с помощью иммуногистохимии (рис. 6А). rM3D(Gs) и mCherry экспрессировались в большинстве дофаминовых нейронов TH + (85,3 ± 4,2% и 87,1 ± 5,3% соответственно), но не экспрессировались в нейронах TH – в ВОП. Экспрессия AAV была преимущественно ограничена VTA и редко наблюдалась в соседних SNc. Таким образом, rM3D(Gs) избирательно экспрессировался в большинстве дофаминовых нейронов VTA.
Хемогенетическое усиление высвобождения дофамина увеличивает самостоятельный прием кокаина у мышей с условным нокаутом Epac2 (Epac2-cKO).
( A ) AAV8-hSyn-DIO-rM3D(Gs)-mCherry (rM3D(Gs)) или AAV8-hSyn-DIO-mCherry (mCherry) билатерально микроинъецировали в вентральную область покрышки (VTA) Epac2-cKO мыши. Иммуногистохимия в срезах VTA показала, что rM3D(Gs) и mCherry экспрессировались в большинстве клеток TH 9.1351 + дофаминовых нейрона, но не в TH — нейрона. ( B ) Слева, репрезентативный сигнал дофамина и временная динамика переходных процессов дофамина в прилежащем ядре (NAc) мышей Epac2-cKO до и после введения дезхлорклозапина (DCZ). Справа, DCZ (10 мкг/кг, внутрибрюшинно) значительно увеличивал частоту переходных процессов дофамина в оболочке NAc мышей Epac2-cKO после предварительной обработки носителем (парный тест t : t 9 = 7,6, p<0,001). ; n=10, 5 мужчин и 5 женщин). ( C ) DCZ не влиял на частоту переходных процессов дофамина у мышей, которым предварительно вводили H89 (1 мг/кг, внутрибрюшинно) (парный тест t-: t 9 =0,1, p=0,889; n=10 , 5 мужчин и 5 женщин). ( D ) Хронология инъекций AAV, самостоятельного приема кокаина и лечения DCZ. ( E ) Предварительная обработка DCZ значительно увеличивала количество активных тычков носом (двусторонний дисперсионный анализ с повторными измерениями: rM3D(Gs), F 1,14 = 12,8, p = 0,003; время, F 2,28 = 1,9, р=0,164; rM3D(Gs) × взаимодействие во времени, F 2,28 = 1,6, p = 0,225) без воздействия на неактивные тычки носом (двусторонний дисперсионный анализ с повторными измерениями: rM3D(Gs), F 1,14 = 0,3, p=0,613, время, F 2,28 = 3,0, p=0,065, rM3D(Gs) × время взаимодействия, F 2,28 = 1,7, p=0,209) Gs)-экспрессирующие мыши Epac2-cKO по схеме с фиксированным соотношением-1 (FR1) (n = 7–9, 4 самца и 3–5 самок). ( Ф ) Предварительное лечение ДКЗ значительно увеличивало количество инъекций кокаина (rM3D(Gs), F 1,14 = 17,8, p<0,001; время, F 2,28 = 3,2, p=0,058; rM3D (Gs) × время взаимодействия, F 2,28 = 1,3, p = 0,283, n = 7–9, 4 самца и 3–5 самок) по схеме FR1 в rM3D(Gs)-экспрессирующем Epac2-cKO мыши. ( G ) DCZ вызывал сдвиг вверх восходящей части кривой доза-реакция кокаина у мышей Epac2-cKO, экспрессирующих rM3D(Gs) (двухфакторный дисперсионный анализ с повторными измерениями: rM3D(Gs), Ж 3,30 = 8,1, р<0,001; доза кокаина, F 6180 = 79,3, p<0,001; rM3D(Gs) × взаимодействие дозы кокаина, F 18 180 = 3,6, p<0,001; n=8–9, 4 мужчин и 4–5 женщин). ns, p>0,05, *p<0,05, **p<0,01, ***p<0,001 (см. также рис. 6 — приложение 1 к рисунку).
В последующих экспериментах мы вводили Cre-зависимый rM3D(Gs)-mCherry в VTA и смесь 1:1 AAV9-hSyn-GRAB 9.1293 DA2m (GRAB DA2m ) и AAV8-hSyn-mCherry (mCherry) в оболочку NAc мышей Epac2-cKO с последующей имплантацией оптического волокна 200 мкм над местом инъекции GRAB DA2m (рис. 6 — дополнение к рисунку). 1А). mCherry использовали в качестве независимого от дофамина контроля изменений флуоресценции, не связанных с активацией GRAB DA2m . После восстановления и экспрессии AAV в течение 2–3 недель была выполнена фотометрия волокон для определения динамики дофамина в NAc. ЗАХВАТИТЬ 9Сенсоры 1293 DA2m представляют собой мутантные рецепторы D 2 , которые увеличивают интенсивность флуоресценции при связывании дофамина и сохраняют чувствительность к антагонистам D 2 (Sun et al., 2020). Мы обнаружили, что транзиенты флуоресценции устранялись внутрибрюшинным введением. инъекция антагониста D 2 галоперидола (0,1 мг/кг, внутрибрюшинно; рисунок 6 — дополнение к рисунку 1B), что указывает на то, что переходные процессы связаны со связыванием дофамина с датчиком.
DCZ является активатором DREADD, который проявляет большее сродство и эффективность в отношении DREADD, чем N-оксид клозапина, обладает высокой проникающей способностью в головной мозг и метаболически стабилен (Nagai et al., 2020). Двухфотонная визуализация in vivo нейронов соматосенсорной коры, коэкспрессирующих G q -DREADD (hM3Dq) и GCaMP6 показали, что i. p. инъекция DCZ приводила к быстрому увеличению активности GCaMP6 у мышей, которая достигала пика примерно через 10 минут и стабилизировалась в течение по крайней мере 150 минут. Тем не менее, мы использовали подкожную инъекцию DCZ, чтобы обеспечить относительно равномерную и длительную активацию Gs- и Gi-DREADD. Подкожное введение DCZ (100 мкг/кг) через 20 мин после внутрибрюшинного введения. Предварительная обработка носителем вызвала быстрое и сильное увеличение (~ 30 минут) частоты переходных процессов дофамина в оболочке NAc, за которым последовало более умеренное и устойчивое увеличение, продолжающееся в течение всего времени записи фотометрии волокон (рис. 6B). Предварительная обработка ингибитором PKA H89(1 мг/кг, внутрибрюшинно) блокировал действие DCZ на повышение частоты переходных процессов дофамина (рис. 6C). Таким образом, DCZ индуцировал PKA-зависимое увеличение высвобождения дофамина у мышей Epac2-cKO.
Мы определили, может ли активация rM3D(Gs) с помощью DCZ в дофаминовых нейронах VTA обратить вспять нарушение самостоятельного приема кокаина у мышей Epac2-cKO. Мыши Epac2-cKO получали двусторонние инъекции Cre-зависимого rM3D(Gs) или mCherry внутри VTA и подвергались катетеризации яремной вены для самостоятельного введения кокаина. График времени инъекции AAV и поведенческих тестов показан на рисунке 6D. Мышей обучали самостоятельному приему стабильного кокаина (0,5 мг/кг/инфузия) в течение 10 дней по схеме FR1. Мы исследовали, влияет ли усиление высвобождения дофамина посредством активации rM3D(Gs) на потребление кокаина у мышей, у которых установилось стабильное самостоятельное введение кокаина. Мышам вводили подкожно DCZ (100 мкг/кг) за 10 минут до тестирования самостоятельного введения кокаина в течение трех ежедневных сеансов, каждый сеанс длился по 3 часа. DCZ увеличил количество активных тычков в нос (рис. 6E) и вливаний кокаина (рис. 6F) у мышей, экспрессирующих rM3D(Gs) Epac2-cKO. Напротив, DCZ не оказывал значительного влияния на активные тычки в нос, неактивные тычки в нос (рис. 6E) или инфузии кокаина (рис. 6F) у мышей Epac2-cKO, экспрессирующих mCherry, что указывает на то, что эффекты DCZ опосредованы активацией rM3D (Gs). . В парадигме самостоятельного приема нескольких доз кокаина DCZ вызывал сдвиг вверх восходящего участка перевернутой U-образной кривой самостоятельного приема кокаина у мышей Epac2-cKO, которые экспрессировали rM3D(Gs) в дофаминовых нейронах VTA, но не изменить кривую зависимости от дозы кокаина у мышей Epac2-cKO, которые экспрессировали контрольный вектор (рис. 6G). Эти результаты позволяют предположить, что активация rM3D(Gs) с помощью DCZ обращала вспять нарушение самостоятельного приема кокаина у мышей Epac2-cKO.
Хемогенетическое ингибирование высвобождения дофамина снижает самостоятельный прием кокаина у мышей дикого типа
Затем мы определили, будет ли селективное ингибирование высвобождения дофамина из дофаминовых нейронов VTA имитировать эффект разрушения Epac2 на самостоятельный прием кокаина у мышей дикого типа. Мыши WT ( Slc6a3 IRES cre/+ ) получали билатеральные интравенозные инъекции AAV8-hSyn-DIO-hM4D(Gi)-mCherry или mCherry и внутри-NAc инъекцию 1:1 смешанного GRAB DA2m и AAV-mCherry с последующей имплантацией оптического волокна 200 мкм (рис. 7А). После выздоровления в течение 2–3 недель высвобождение дофамина контролировали с помощью фотометрии волокон in vivo. Подкожная инъекция DCZ (100 мкг/кг) значительно снижала частоту транзиентов дофамина в оболочке NAc мышей, которые экспрессировали hM4D(Gi), но не mCherry (рис. 7B). После завершения экспериментов экспрессию AAV в VTA и NAc подтверждали с помощью иммуногистохимии. hM4D(Gi) и mCherry экспрессировались в большинстве TH + дофаминовых нейрона (83,8 ± 4,1% и 84,7 ± 3,4% соответственно), но не экспрессировались в нейронах TH — в VTA (рис. 7А). Таким образом, hM4D(Gi) избирательно экспрессировался в большинстве дофаминовых нейронов VTA.
Хемогенетическое ингибирование высвобождения дофамина снижает самостоятельный прием кокаина у мышей дикого типа (WT).
( A ) AAV8-hSyn-DIO-hM4D(Gi)-mCherry (hM4D(Gi)) или AAV8-hSyn-DIO-mCherry (mCherry) билатерально микроинъецировали в вентральную область покрышки (VTA) мышей дикого типа, и дофаминовый сенсор AAV9-hsyn-DA2m (GRAB DA2m ) вводили в оболочку NAc с последующей имплантацией волоконно-оптической канюли. Иммуногистохимия показала, что hM4D(Gi) экспрессировался в большинстве дофаминовых нейронов TH + (зеленый), но не экспрессировался в нейронах TH — в VTA. ( B ) Слева, репрезентативный сигнал дофамина в прилежащем ядре (NAc) мышей дикого типа до и после лечения дезхлорклозапином (DCZ). Справа, DCZ (10 мкг/кг, внутрибрюшинно) не изменял частоту дофаминовых транзиентов у мышей с экспрессией mCherry (парный тест t : t 8 = 0,7, p = 0,514; n = 9, 5). мужчин и 4 женщин). Частота дофаминовых транзиентов была значительно снижена после обработки DCZ у мышей дикого типа, экспрессирующих hM4D(Gi) в дофаминовых нейронах VTA (спаренные t -тест: t 9 =2,9, р=0,017; n=10, 5 мужчин и 5 женщин). ( C ) Хронология самостоятельного приема кокаина и лечения DCZ. ( D–E ) Предварительное лечение DCZ значительно уменьшило активные тычки в нос (двухфакторный анализ с повторными измерениями: hM4D(Gi), F 1,15 = 18,6, p<0,001; время, F 2 ,30 = 0,03, p = 0,966; hM4D(Gi) × время взаимодействия, F 2,30 = 2,5, p = 0,096) и инфузии кокаина (двухфакторный дисперсионный анализ с повторными измерениями: hM4D(Gi), Ж 1,15 = 25,7, р<0,001; время, Ж 2,30 = 2,2, р=0,132; hM4D(Gi) × время взаимодействия, F 2,30 = 0,02, p=0,986) без изменения неактивных тычков носом (hM4D(Gi), F 1,15 = 0,01, p=0,911; время , F 2,30 = 0,5, p=0,627; =8–9, 4 мужчин и 4–5 женщин). ( F ) DCZ вызывал значительный сдвиг кривой доза-реакция кокаина вниз (двусторонний дисперсионный анализ с повторными измерениями: hM4D(Gi), Ж 3,26 = 13,8, р<0,001; доза кокаина, F 6156 = 100,2, p<0,001; hM4D(Gi) × взаимодействие дозы кокаина, F 18 156 = 2,2, p = 0,006; n = 7–8, 4 самца и 3–4 самки) у мышей WT, которые экспрессировали hM4D (Gi) в дофаминовых нейронах VTA. нс, р>0,05, *р<0,05, **р<0,01, ***р<0,001.
Показав, что активация hM4D(Gi) снижает высвобождение дофамина, мы затем исследовали ее влияние на самостоятельный прием кокаина у мышей дикого типа. График времени инъекции AAV и поведенческих тестов показан на рисунке 7C. Мышей обучали самостоятельному приему стабильного кокаина (0,5 мг/кг/инфузия) в течение 10 дней по схеме FR1. Мыши продолжали принимать кокаин самостоятельно в последующие дни, получая подкожные инъекции DCZ (100 мкг/кг) за 10 минут до тестирования. DCZ значительно уменьшил количество активных уколов в нос и вливаний кокаина у мышей WT, экспрессирующих hM4D(Gi), но не оказал значительного влияния на активные уколы в нос, вливания кокаина или неактивные уколы в нос у мышей WT, экспрессирующих mCherry (рис. 7D-E). . В парадигме самостоятельного приема нескольких доз кокаина DCZ приводил к сдвигу вниз кривой доза-реакция кокаина у мышей WT, которые экспрессировали hM4D(Gi) в дофаминовых нейронах VTA, но не изменял кривую доза-реакция кокаина у мышей WT, которые выраженный mCherry (рис. 7F). Эти результаты показывают, что опосредованное hM4D(Gi) ингибирование высвобождения дофамина снижает самостоятельный прием кокаина и имитирует эффект разрушения Epac2.
Обсуждение
Здесь мы показываем, что ингибирование Epac2 или генетическая делеция в дофаминовых нейронах среднего мозга уменьшали самостоятельный прием кокаина при многократном приеме кокаина и схемах подкрепления. Epac2-cKO приводил к снижению вызванного высвобождения дофамина, тогда как агонизм Epac2 усиливал высвобождение дофамина в NAc. Чтобы определить, способствует ли ослабленный дофаминовый ответ нарушению самостоятельного приема кокаина, мы селективно экспрессировали rM3D(Gs) или hM4D(Gi) в дофаминовых нейронах VTA и обнаружили, что усиление высвобождения дофамина с активацией rM3D(Gs) обращало вспять снижение кокаина. самостоятельный прием у мышей Epac2-cKO, в то время как снижение высвобождения дофамина с активацией hM4D(Gi) ослабляло самостоятельный прием кокаина у мышей WT. Гипотетическая модель, обобщающая эти результаты, показана на рисунке 8. В совокупности эти результаты показывают критическую функцию Epac2 в модуляции высвобождения дофамина и кокаинового подкрепления.
Гипотетическая модель, объясняющая взаимосвязь между нарушением работы Epac2, снижением высвобождения дофамина и нарушением самостоятельного приема кокаина.
( A ) Известно, что Epac2 взаимодействует с белками активной зоны RIM, Munc13 и Rab3. Потенциальное взаимодействие Epac2 с белками активной зоны в варикозном расширении аксонов дофаминовых нейронов способствует экзоцитозу дофамина, который поддерживает кокаиновое подкрепление во время самостоятельного введения (SA). ( B ) Мыши с условным нокаутом Epac2 (Epac2-cKO) приводили к снижению высвобождения дофамина и кокаиновой SA. ( C ) hM4D(Gi)-опосредованное ингибирование запуска потенциала действия (AP) и высвобождение дофамина из аксональных окончаний дофаминовых нейронов вентральной области покрышки (VTA) у мышей дикого типа приводило к снижению самостоятельного приема кокаина (имитируя эффект Epac2-cKO). ( D ) Усиление высвобождения дофамина за счет активации rM3D(Gs) в дофаминовых нейронах VTA увеличивает высвобождение дофамина через путь PKA и усиливает самостоятельный прием кокаина у мышей Epac2-cKO.
ESI-05 вызывал дозозависимое снижение потребления кокаина у мышей дикого типа при схеме подкрепления FR1. В парадигме самостоятельного введения нескольких доз ESI-05 дозозависимо снижал инфузии кокаина и потребление в дозах кокаина, которые попадают как на восходящую, так и на нисходящую ветви перевернутой U-образной кривой доза-реакция. Точно так же, хотя мышам Epac2-cKO и WT требовалось одинаковое количество тренировок для достижения стабильного самостоятельного приема кокаина, мыши Epac2-cKO продемонстрировали общее снижение самостоятельного приема при низких и высоких стандартных дозах кокаина. Это снижение кривой доза-реакция предполагает общее снижение мотивации к самостоятельному приему кокаина, а не простое изменение активности кокаина (Mello and Negus, 19).96). В подтверждение этого, ингибирование Epac2 или генетическая делеция также уменьшали активные тычки носом, вливания кокаина и точку останова при подкреплении PR. ESI-05 и Epac2-cKO не влияли на неактивные тычки в нос, а также не влияли на самостоятельный пероральный прием сахарозы при схемах подкрепления FR1 или PR. Таким образом, снижение уровня самостоятельного приема кокаина вряд ли было вызвано общим подавлением оперантных реакций. Вместе эти исследования показывают, что Epac2 в дофаминовых нейронах среднего мозга вносит решающий вклад в подкрепление кокаина во многих парадигмах самостоятельного приема.
Кокаин и сахароза повышают уровень дофамина, уменьшая его поглощение и увеличивая его высвобождение, соответственно (Anderson and Pierce, 2005; Patriarchi et al., 2018). Было несколько неожиданно, что ESI-05 и Epac2 ослабляли самостоятельный прием кокаина, но существенно не изменяли самостоятельный прием сахарозы. Однако сообщалось об аналогичной дифференциальной модуляции самостоятельного приема кокаина и сахарозы фармакологическими агентами (Levy et al., 2007; Romieu et al., 2008; Mu et al., 2021). Хотя в нейронных цепях, кодирующих вознаграждение за еду и наркотики, существует значительное перекрытие (Volkow et al., 2012), большинство нейронов NAc демонстрировали разные паттерны возбуждения в ответ на вознаграждение за еду и кокаин (Carelli, 2002). Как упоминалось во введении, повторное употребление кокаина приводит к снижению G 9экспрессия 1293 αi/o в VTA и усиление передачи сигналов цАМФ (Nestler et al. , 1990; Striplin and Kalivas, 1992). Вполне вероятно, что Epac2 преимущественно активируется после хронического самостоятельного приема кокаина, что может объяснить, почему нарушение Epac2 ослабляет самостоятельный прием кокаина, но не сахарозы.
Было показано, что агонизм Epac способствует экзоцитозу везикул путем взаимодействия с белками активной зоны RIM и Munc13 (Ozaki et al., 2000; Tengholm and Gylfe, 2017). RIM сгруппирован в дофаминовых аксонах, и условное нокаутирование RIM из дофаминовых нейронов среднего мозга отменяет высвобождение дофамина, запускаемое потенциалом действия (Liu and Kaeser, 2019).). Мы проверили гипотезу о том, что Epac2 модулирует высвобождение дофамина аксонами в стриатуме. Используя FSCV для регистрации вызванного высвобождения дофамина в срезах головного мозга, мы обнаружили, что Epac2-селективный агонист дозозависимо увеличивает вызванное высвобождение дофамина в NAc. Этот эффект устранялся ингибированием Epac2 и генетической делецией Epac2 , но на него не влияло ингибирование PKA. Кроме того, у мышей Epac2-cKO наблюдалось ослабление вызванного высвобождения дофамина in vitro в NAc и дорсальном стриатуме. RIM опосредует связывание пузырьков и необходим для Ca 2+ — канальный набор в места выпуска (Acuna et al., 2016). Следовательно, Epac2 может способствовать экзоцитозу дофамина из дофаминовых нейронов среднего мозга путем взаимодействия с RIM и др. белками активной зоны и вызывая Ca 2+ -зависимое высвобождение дофамина.
Существует вероятность того, что агонист Epac2 S-220 влияет на другие системы нейротрансмиттеров, косвенно модулируя высвобождение дофамина. Оптогенетическая стимуляция холинергических интернейронов в NAc индуцировала высвобождение дофамина посредством активации никотиновых ацетилхолиновых рецепторов (nAChR) (Cachope et al., 2012; Threlfell et al., 2012). ГАМК тонически подавляет высвобождение дофамина аксонами в дорсальном стриатуме через ГАМК 9Рецепторы 1293 A и ГАМК B (Roberts et al., 2020). Таким образом, ацетилхолин и ГАМК оказывают косвенное, но существенное влияние на выброс дофамина. Однако S-220-индуцированное усиление высвобождения дофамина устранялось Epac2-cKO из дофаминовых нейронов, что позволяет предположить, что дофаминовые окончания аксонов являются преобладающим местом действия S-220. В будущих исследованиях было бы интересно проверить, изменяется ли S-220-индуцированное усиление высвобождения дофамина под действием блокаторов рецепторов нАХР и ГАМК.
Злоупотребление наркотиками, включая кокаин, повышает уровень дофамина в NAc, что, как считается, лежит в основе их подкрепляющего действия (Anderson and Pierce, 2005; Volkow et al., 2006). Поскольку нарушение Epac2 приводит к снижению синаптически высвобождаемого дофамина, вполне вероятно, что опосредованное кокаином ингибирование DAT у мышей с нарушенным Epac2 будет вызывать меньшее повышение внеклеточных уровней дофамина и ослабление поведения, обусловленного дофамином. Чтобы проверить, способствует ли снижение высвобождения дофамина нарушению самостоятельного приема кокаина, мы экспрессировали rM3D(Gs) в дофаминовых нейронах VTA у мышей Epac2-cKO и обнаружили, что DCZ увеличивает частоту переходных процессов дофамина за счет активации PKA и увеличения приема кокаина при самостоятельном введении. Армирование FR1. В парадигме самостоятельного введения нескольких доз активация rM3D (Gs) вызывала сдвиг вверх восходящей ветви кривой доза-реакция кокаина, не затрагивая нисходящей ветви. Увеличение оперантной реакции на восходящую ветвь может быть связано с усилением подкрепляющей эффективности кокаина (Mello and Negus, 19).96; Кейн и др., 2002; Робертс и др., 2013). Поскольку активация rM3D(Gs) увеличивает высвобождение дофамина, инфузии кокаина в малых дозах, вероятно, приводят к повышению уровня дофамина, что усиливает подкрепление и способствует оперантному реагированию (Mello and Negus, 1996; Caine et al., 2002; Roberts et al., 2013). ). DCZ-индуцированное увеличение подкрепляющей эффективности кокаина может противодействовать эффекту снижения скорости более высоких доз кокаина (Zimmer et al., 2011), что может объяснить, почему активация rM3D(Gs) не вызывала значительного смещения нисходящей конечности. Вместе эти эксперименты демонстрируют, что нарушение самостоятельного приема кокаина, вызванное разрушением Epac2, в значительной степени обратимо при восстановлении высвобождения дофамина в среднем мозге.
Напротив, активация hM4D(Gi) у мышей WT ( Slc6a3 IRES cre/+ /Epac2 wt/wt ) снижала частоту переходных процессов дофамина in vivo, NAc самостоятельный прием кокаина по схеме подкрепления FR1 и вызывал общий сдвиг кривой доза-реакция кокаина вниз. Этот фенотип отражает фенотип мышей Epac2-cKO и указывает на снижение самоконтроля во всем диапазоне стандартных доз кокаина (Mello and Negus, 19).96). Точно так же хемогенетическое ингибирование дофаминовых нейронов VTA посредством hM4D(Gi) подавляло самостоятельный прием героина по схеме подкрепления FR1 (Corre et al., 2018). И наоборот, нокдаун ауторецепторов D2 в дофаминовых нейронах среднего мозга повышал уровень дофамина из-за снижения аутоингибирования и вызывал общий сдвиг кривой доза-реакция кокаина вверх (Chen et al., 2018). Таким образом, двунаправленная модуляция высвобождения дофамина способна усиливать или подавлять оперантный ответ на кокаин при множественных стандартных дозах и не приводит к простому изменению активности кокаина. Кроме того, Gq-DREADD-опосредованная стимуляция дофаминовых нейронов VTA потенцировала восстановление, вызванное сигналом и кокаином, тогда как Gi-DREADD уменьшало восстановление (Mahler et al., 2018).
Мы признаем ограничения наших экспериментов DREADD. Во-первых, стимуляция Gs-DREADD использовалась для восстановления высвобождения дофамина у мышей Epac2-cKO, но не восстанавливала функцию Epac2. Дальнейшие исследования могли бы определить, может ли вирусная экспрессия Epac2 в дофаминовых нейронах VTA у мышей Epac2-cKO восстановить высвобождение дофамина и самостоятельный прием кокаина до уровня мышей WT. Во-вторых, наши исследования DREADD были ограничены экспериментами, которые были необходимы для проверки центральной гипотезы о том, что Epac2-cKO-индуцированное снижение высвобождения дофамина способствует дефициту самостоятельного приема кокаина. И мимикрия, и окклюзия могут дать представление о том, происходят ли два биологических процесса по одному и тому же механизму. Мы показали, что ингибирование hM4D(Gi) дофаминовых нейронов VTA у мышей WT имитировало эффекты Epac2-cKO при самостоятельном приеме кокаина. Дополнительный эксперимент должен был бы определить, снижает ли ингибирование hM4D(Gi) дофаминовых нейронов дальнейшее снижение самостоятельного введения кокаина у мышей Epac2-cKO или эффект ингибирования hM4D(Gi) блокируется Epac-cKO.
Вполне вероятно, что Epac2 регулирует поведение с помощью механизмов, выходящих за рамки облегчения пресинаптического высвобождения дофамина. Epac передает сигналы вниз по течению через множество эффекторов, включая Rap, Ras, Rho, фосфолипазу C и митоген-активируемые протеинкиназы (Laurent et al., 2012). Epac может активироваться как цАМФ-зависимыми, так и независимыми механизмами (Shibasaki et al., 2004; Baameur et al., 2016; Robichaux and Cheng, 2018). Повторное введение кокаина индуцирует структурную пластичность NAc (Cahill et al., 2016; Barrientos et al., 2018; Kalivas, 2018), в которой ключевую роль играет передача сигналов Rap1 GTPase (Cahill et al. , 2016). Самостоятельный прием и поиск кокаина зависят как от дофамин-зависимых, так и от -независимых механизмов (Schmidt and Pierce, 2010; Mereu et al., 2020). Важной функцией Epac является регулирование синаптической передачи посредством как пресинаптических, так и постсинаптических механизмов. Постсинаптически Epac2 модулирует транспортировку AMPAR, индуцируя изменения в составе субъединиц AMPAR (Woolfrey et al., 2009).; Liu et al., 2016) и увеличивает вероятность открытого канала GluA3-AMPAR (Gutierrez-Castellanos et al., 2017). Пресинаптически Epac2 способствует высвобождению глутамата и обеспечивает долговременную потенциацию (LTP) (Fernandes et al., 2015; Martín et al., 2020). Генетическая делеция Epac1/2 или Epac2 приводит к нарушению LTP в гиппокампе, пространственному социальному взаимодействию, пространственному обучению и памяти (Yang et al., 2012). Наркотики, вызывающие привыкание, такие как кокаин, вызывают долговременную синаптическую пластичность в схеме вознаграждения мозга (Wolf, 2016; Dong et al. , 2017; Wang et al., 2018; Zinsmaier et al., 2022). Мы показали, что Epac2, но не Epac1, необходим для изменений возбуждающей и тормозной передачи в дофаминовых нейронах VTA, вызванных непроизвольным воздействием кокаина (Liu et al., 2016; Tong et al., 2017). Вполне вероятно, что Epac2 действует через многофакторные механизмы, которые вместе способствуют нейроадаптации, вызванной кокаином, и подкреплению наркотиками.
Материалы и методы
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information | ||
|---|---|---|---|---|---|---|
| Strain, strain background ( Mus musculus ) | C57BL/6J mice | Лаборатория Джексона | Товарный №: 000664 | |||
| Генетический реагент ( Mus musculus ) | Гетерозиготный SLC6A3 IRES CRE/+ Мыши | Вирус ) | AAV9-HSYN-GRAB DA2M | WZ Biosciences | ||
| штамм, фон штамма ( Adeno-Associated ) | Aav82-HSYN-MCHRIN-MCHRIN-MCHRIN-MCHRIN-MCHRIN-MCHERS8. 2085 | Addgene | Catalog#: 114472-AAV8 | |||
| Strain, strain background ( Adeno-associated virus ) | AAV8-hSyn-DIO-rM3D(Gs)-mCherry | Addgene | Catalog#: 50458-AAV8 | |||
| Strain, strain background ( Adeno-associated virus ) | AAV8-hSyn-DIO-hM4D(Gi)-mCherry | Addgene | Catalog#: 44362-AAV8 | |||
| Напряжение, фон напряжения ( Адено-ассоциированный вирус ) | AAV8-HSYN-Dio-Mcherry | ADDGENE | КАТАЛОГ №: 50459-AAV8 | |||
| Antibode | Rablonli-Thatei-Thth-Thth-Thate-Thath. , Lot: C2707 | 1:300 | ||||
| Antibody | Goat anti-rabbit IgG Alexa Fluor-488 | Cell Signaling | Stock#: 4412 | 1:300 | ||
| Other | NeuroTrace 640/ 660 темно-красный флуоресцентный краситель Ниссля | Thermo Fisher Scientific | N-21483 | 1:150 | ||
| Software, algorithm | Origin 2022b | OriginLab Corporation | ||||
| Software, algorithm | RStudio | RStudio, PBC | Ссылка на DIO (Github): GitHub — xiaojieliu17/Fiber-photometry |
Животные
Содержание и использование животных осуществлялись в соответствии с протоколами, утвержденными Институциональным комитетом по уходу и использованию животных Медицинского колледжа штата Висконсин. гомозиготный 9Мышей 0004 Epac2 -floxed ( Epac2 loxP/loxP ) получали, как описано ранее (Pereira et al., 2013). Гетерозиготных мышей Slc6a3 IRES cre/+ (номер Jax: 006660) получали из лаборатории Джексона (Бар-Харбор, Мэн). Все линии мышей поддерживали на одном и том же фоне C57BL/6. Мыши, нокаутированные по дофаминовым нейронам Epac2 (Epac2-cKO), были получены путем скрещивания гомозиготных мышей Epac2 loxP/loxP с гетерозиготными Slc6a3 IRES cre/+ мыши. Затем мы пересекли эти соединения SLC6A3 IRES CRE/+ /EPAC2 LOXP/WT мышей EPAC2 LOXP/LOXP с использованием мыши DOPAMINE Necroin Specific-Specifice-Specifice Specifice Specific-Specifice Specifice-Specific-Specifice-Specifice-Specififie-Specififie-Specififie Necroin-Specific-Specific-Specifice-Specific. IRES cre/+ /Epac2 loxP/loxP ). Slc6a3 IRES cre/+ /Epac2 вес/вес или 9В качестве группы WT использовали мышей 0004 Slc6a3 +/+ /Epac2 loxP/loxP мышей. Генотипирование проводили с помощью ПЦР-анализа биоптатов уха или хвоста.
Всем животным предоставлялся свободный доступ к пище и воде в помещении с контролируемой температурой (23 ± 1°C) и влажностью (40–60%) при обратном световом цикле. Для экспериментов использовали взрослых мышей обоего пола (10–12 недель). Мышей обрабатывали ежедневно в течение 3–6 дней до экспериментов. Мышей помещали в одиночное помещение после внутривенной катетеризации и/или имплантации волоконно-оптической канюли, чтобы предотвратить повреждение имплантированных катетеров и канюль соседями по клетке.
Стереотаксические операции и инъекции ААВ
Запросить подробный протокол
Мышей анестезировали кетамином (90 мг/кг, внутрибрюшинно) и ксилазином (10 мг/кг, внутрибрюшинно) и помещали в роботизированную стереотаксическую систему (Neurostar, Тюбинген, Германия). Объем AAV (см. ниже) вводили через стеклянный капиллярный наноинжектор (Neurostar) или программируемый нанолитровый инжектор Nanoject III (Drummond Scientific Company, Broomall, PA). Скорость введения составляла 60 нл/мин, а инжектор оставался на месте в течение 5 мин для обеспечения адекватной диффузии от наконечника инжектора (~30 мкм). Мышей держали на грелке во время операции и в течение 2–3 часов после операции для поддержания температуры тела. После операции животным вводили подкожно анальгетик (бупренорфин-SR, 0,05 мг/кг) и давали возможность восстановиться примерно за 2 недели до проведения поведенческих тестов (см. ниже).
Для экспериментов, в которых изучалась активация DREADD в дофаминовых нейронах VTA при высвобождении дофамина в NAc, AAV8-hSyn-DIO-rM3D(Gs)-mCherry, AAV8-hSyn-DIO-hM4D(Gi)-mCherry или AAV8-hSyn- DIO-mCherry (по 200 нл каждый; Addgene, Watertown, MA) билатерально вводили в VTA либо мышам дикого типа ( Slc6a3 IRES cre/+ /Epac2 wt/wt ), либо мышам (Epac2-cKO АП, –3,1 мм, ML, ±1,0 мм, DV, –4,8 мм (угол 7°). Затем 200 нл смеси 1:1 AAV9-hSyn-GRAB DA2m и AAV8-hSyn-mCherry вводили в оболочку NAc. После инъекции AAV волоконно-оптическую канюлю с низкой аутофлуоресценцией (OD = 200 мкм, NA = 0,57; Doric Inc, Квебек, Канада) опускали в NAc чуть выше места инъекции AAV. Канюля была закреплена в черепе двумя маленькими винтами (00–96 × 1/16; Invivo1), которые были прикреплены к черепу быстроклеящимся цементом C&B Metabond.
Для экспериментов, в которых изучалось влияние активации DREADD в дофаминовых нейронах VTA на самостоятельный прием кокаина, мышам сначала подвергали катетеризацию яремной вены (см. ниже), затем AAV8-hSyn-DIO-rM3D(Gs)-mCherry, AAV8-hSyn- DIO-hM4D(Gi)-mCherry или AAV8-hSyn-DIO-mCherry билатерально вводили в VTA мышам Epac2-cKO или WT (9).0004 Slc6a3 IRES cre/+ /Epac2 вес/вес ), как описано выше.
Катетеризация яремной вены
Запросить подробный протокол
Мышей анестезировали кетамином (90 мг/кг, внутрибрюшинно) и ксилазином (10 мг/кг, внутрибрюшинно) и вводили полиуретановый катетер с закругленным концом с выступом на расстоянии 1,2 см от кончика катетера (#C20PU-MJV1934, Instech Laboratories, Inc, Plymouth Meeting, PA) вводили в правую яремную вену. Катетер присоединяли к кнопке сосудистого доступа (размер 25; #VAM1B/25, Instech) и имплантировали подкожно на спину мышам. Эта система уменьшает травмирование внутренней оболочки стенок сосудов и позволяет поддерживать положительное давление, что значительно продлевает проходимость катетера и позволяет проводить групповое размещение (Slosky et al., 2020; Slosky et al., 2021). Катетеры промывали 0,1 мл гепаринизированного солевого раствора (30 ЕД/мл) после операции для предотвращения коагуляции. Тесты на проходимость катетера (ксилазин, 2 мг/кг, 0,5 мл, в/в . ) выполнялись еженедельно после самостоятельного введения или при подозрении на повреждение катетера. Животные, не отвечающие критериям проходимости (седативное действие в течение 5 с), исключались.
Самостоятельный прием кокаина по схемам FR1 и PR
Запросить подробный протокол
Через 5–7 дней после хирургического восстановления мышей обучали самостоятельному приему кокаина в течение 3-часовых сеансов в камерах оперантного кондиционирования (Med. Associates, St Albans, VT). Положение активного и неактивного носовых портов было уравновешено. Каждый ответ на активный операндум приводил к вливанию кокаина (т. е. по схеме FR1; 40–50 мкл в течение 2,3–2,8, в зависимости от веса мышей) и освещению сигнального индикатора над активным носовым портом на 5 с с последующим тайм-аут 10 с. Ответы во время тайм-аутов учитывались, но не имели последствий. Сессии заканчивались раньше, если было заработано 64 поощрения, чтобы избежать передозировки кокаином. Каждая активная инъекция в нос доставляла 0,5 мг/кг/инфузию кокаина в течение 10 дней, пока не было достигнуто стабильное самостоятельное введение. Стабильный самостоятельный прием определяли как: ≥25 инфузий и соотношение активного/неактивного ответа >2:1 в течение 3 дней подряд и менее 20% вариабельности ежедневных инфузий препарата в течение двух последовательных сеансов. Мышей, которые не достигли критериев после 10 дней обучения, исключали из дальнейшего исследования. После завершения 10-дневного обучения самостоятельному введению мышам вводили ESI-05 (3,3 или 10 мг/кг внутрибрюшинно) или носитель (0 мг/кг, внутрибрюшинно) за 10 минут до тестирования самостоятельного введения при подкреплении FR1 в дизайн латинского квадрата. Для и.п. инъекций, ESI-05 сначала растворяли в ДМСО для получения исходного раствора с концентрацией 30 мг/мл, делили на аликвоты и хранили в морозильной камере при –80°C. Перед использованием исходный раствор сначала смешивали с таким же объемом Tween-80, а затем разбавляли физиологическим раствором (0,9% NaCl) для получения рабочего раствора (ДМСО:Твин-80:физиологический раствор = раствор 1:1:18). Тот же раствор использовали в качестве контроля транспортного средства. После медикаментозного лечения мыши, как и раньше, сами принимали кокаин до тех пор, пока реакция не вернулась к стабильным уровням, используя вышеуказанные критерии. Последующие дозы ESI-05 или носителя затем тестировали в дизайне внутри субъектов.
Дополнительные группы мышей обучали по графику PR. После первого установления стабильного самостоятельного приема кокаина по первоначальной 10-дневной схеме FR1 мышей переводили на схему PR с использованием кокаина в дозе 0,5 мг/кг. График PR требует, чтобы субъект делал все большее количество оперантных ответов (тычки носом) на каждое последующее вознаграждение (вливание кокаина). В графике PR использовались серии 1, 1, 2, 4, 5, 7, 10, 13, 16, 20, 25, 31, 38, 47, 57, 69., 84, 102, … по уравнению: Ν=5 (e вливание × 0,18 –1) (Richardson and Roberts, 1996). Точка останова определяется как максимальное количество нажатий рычага, выполненных для последней инфузии кокаина до 1-часового периода, в течение которого не было получено ни одной инфузии. Когда контрольная точка PR стабилизировалась в течение 3 дней подряд (определяется как вариабельность количества инфузий <15% за этот период), мыши получали ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) или носитель (0 мг/кг, внутрибрюшинно) 10 мин до тестирования на самоуправление кокаином. После медикаментозного лечения мыши сами принимали кокаин, как и раньше, до тех пор, пока реакция не возвращалась к стабильным уровням, а затем были протестированы последующие дозы ESI-05 или носителя в дизайне внутри субъектов.
Чтобы проверить влияние генетической делеции Epac2 в дофаминовых нейронах на самостоятельный прием кокаина, мышей Epac2-cKO и WT сначала обучали самостоятельному приему кокаина по схеме FR1, как описано выше. После 10 дней самостоятельного введения кокаина FR1 мышей переводили на график подкрепления PR (0,5 мг/кг/инфузия). Активные тычки в нос, неактивные тычки в нос и инфузии кокаина по схеме FR1, а также инфузии кокаина и точки останова по схеме PR сравнивали между мышами WT и Epac2-cKO.
Самостоятельный прием нескольких доз кокаина
Запросить подробный протокол
Сначала мышей обучали самостоятельно принимать кокаин по схеме FR1, как описано выше. После того, как были соблюдены критерии стабильного поддерживаемого ответа на кокаин, мышей переводили на самостоятельное введение нескольких доз кокаина, поддерживаемое полным диапазоном доз кокаина (см. Ниже) в сеансе ответа на одну дозу по схеме подкрепления FR1. Тренировочные занятия по схеме многократного приема начинались с 30-минутного приема тренировочной дозы кокаина (0,5 мг/кг/инфузия, продолжительность 3,53 с), затем проводились еще шесть 30-минутных испытаний, в которых различные дозы кокаина предъявлялись в порядке убывания (Anderson et al. , 2018) (1,0, 0,25, 0,125, 0,0625, 0,03125 и 0,0 мг/кг/инфузия; продолжительность инфузии составляла 7,07, 1,77, 0,88, 0,44, 0,22, 0 с соответственно), испытание каждой дозы с 1-минутным перерывом между испытаниями. Предварительное тестирование ESI-05 начиналось после того, как стабилизировалось поведение, связанное с самостоятельным введением кокаина, и стабильность определялась как: (1) менее чем 20-процентное изменение общего количества вливаний кокаина в течение двух последовательных тестовых сессий; (2) доза кокаина, которая поддерживала максимальную частоту ответов, варьировалась не более чем на половину логарифмической единицы в течение двух последовательных сеансов тестирования (Keck et al., 2014; Xi et al., 2017). Мыши получали носитель (0 мг/кг) или ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) за 10 минут до сеанса тестирования, и порядок испытаний для различных доз ESI-05 или носителя был уравновешен. После обработки каждым носителем или ESI-05 мышам давали возможность самостоятельно принимать кокаин в соответствии со схемой многократных доз до тех пор, пока реакция не стабилизировалась, как определено выше, прежде чем последующие дозы ESI-05 или носителя были протестированы в дизайне внутри субъектов. Мышей Epac2-cKO и WT также обучали самостоятельно принимать кокаин с использованием этого протокола, и результаты сравнивали между группами.
Пероральное самостоятельное введение сахарозы
Запросить подробный протокол
мыши C57BL/6 были помещены в условия обратного светового цикла без ограничений в еде и подвергались обработке в течение 3–6 дней до процедур. Эксперименты начинались с ежедневных 1-часовых тренировок в течение 10 дней без какой-либо предварительной оперантной подготовки. Каждый ответ на активный операндум приводит к доставке сахарозы (10% сахарозы; график FR1; 35 мкл в течение 1,98 с) и включению светового индикатора (над носовым портом) в течение всего 5 с и 10-секундного тайм-аута. Ответы во время тайм-аута учитывались, но не имели последствий. Сессии заканчивались раньше, если было заработано 64 подкрепления. Стабильный самостоятельный прием сахарозы определялся как: ≥30 инфузий и соотношение активного/неактивного ответа >2:1 в течение 4 дней подряд, включая последний день, и вариабельность ежедневных инфузий менее 20% в течение двух последовательных сеансов. Животные, не достигшие критериев после 10 дней обучения, исключались из дальнейшего исследования. После завершения 10-дневного обучения самостоятельному введению мышам вводили ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) или носитель (0 мг/кг, внутрибрюшинно) за 10 минут до тестирования на самостоятельное введение в условиях FR1. график в стиле латинского квадрата. После лекарственного лечения животные, как и раньше, самостоятельно вводили сахарозу до тех пор, пока реакция не возвращалась к стабильным уровням, используя вышеуказанные критерии. Последующие дозы ESI-05 или носителя затем тестировали в дизайне внутри субъектов. Дополнительные группы мышей первоначально обучали по схеме подкрепления FR1. После установления стабильного подкрепления FR1 животных переводили на схему подкрепления PR. Когда контрольная точка PR стабилизировалась в течение 3 дней подряд (определялась как вариабельность количества инфузий <15% за этот период), мыши получали инъекцию ESI-05 (3,3 или 10 мг/кг, внутрибрюшинно) или инъекцию носителя за 10 минут до самостоятельного введения сахарозы. тестирование в дизайне латинского квадрата.
Мышей Epac2-cKO и WT обучали самостоятельному введению сахарозы по схеме FR1 в течение 10 дней. После обучения самостоятельному введению сахарозы FR1 мышей переводили на график PR на 3 дня. Между группами сравнивали активные и неактивные тычки в нос, потребление сахарозы и контрольную точку PR.
Тест двигательной активности
Запросить подробный протокол
Для оценки базальной локомоции у мышей WT и Epac2-cKO использовали изготовленную на заказ камеру открытого поля (длина 40 см × ширина 40 см × глубина 40 см). Мышам позволяли свободно исследовать арену в течение 60-минутного тестового сеанса. Двигательную активность регистрировали с помощью автоматизированной системы видео-слежения (ANY-maze; Stoelting, Wood Dale, IL). Расстояние, пройденное в камере, регистрировали и рассчитывали с 5-минутными интервалами.
Фотометрия волокна in vivo для обнаружения высвобождения дофамина в NAc с помощью GRAB
DA2m
Запросить подробный протокол
Для мониторинга флуоресценции дофамин-зависимых GRAB DA2m и дофамин-независимых репортеров mCherry в NAc ведущих мышей использовали двухволновую фотометрическую систему (Doric Lenses, Inc. ) (Liu et al., 2022). Вкратце, управляемая компьютером консоль фотометрии волокон и маломощные драйверы светодиодов (LEDD_4) генерировали возбуждающий свет от двух светодиодов (465 и 560 нм). Блокирующий усилитель синусоидально модулировал интенсивность светодиодов на несинхронных частотах (572,21 и 1017 Гц для 465 и 560 нм соответственно) для фильтрации потенциальной флуоресценции, возникающей из-за перекрытия спектров возбуждения репортеров. Флуоресцентный MiniCube (Doric; FMC6_IE(400-410)_E(460-490)_F(500-550)_ E(555-570)_F(580-680)_S) комбинировал и направлял свет возбуждения через волоконно-оптический патч-корд с низкой автофлуоресценцией (Doric; MFP_200/230/900–0,57_1 .0_FCM_ZF1.25_LAF) и вращающееся соединение (Doric; FRJ_200/230/LWMJ-0.57_1m_FCM_0.15_FCM). Патч-корд был соединен с имплантированной оптоволоконной канюлей через ответную втулку 1,25 мм (Doric; 1,25 мм с черной крышкой; Sleeve_ZR_1,25-BK). Выбросы флуоресценции возвращались через тот же коммутационный шнур и MiniCube, разделялись и направлялись на фотоприемники (модель 2151, низкое значение постоянного тока; Newport Corporation, Ирвин, Калифорния).
Перед каждым экспериментом патч-корд подвергали фотообесцвечиванию возбуждающим светом с длиной волны 465 и 560 нм для дальнейшего уменьшения аутофлуоресценции оптоволокна. Окончательная выходная мощность на конце патч-корда была доведена до 20 мкВт перед записью. Сбор данных контролировался программным обеспечением Doric Neuroscience Studio со скоростью 12,0 ksps, прореженной в 100 раз. Результирующие сигналы демодулировались и подвергались низкочастотной фильтрации с частотой 6 Гц. После прикрепления патч-корда мышей помещали в камеру открытого поля не менее чем на 15 минут для привыкания к окружающей среде перед началом записи. Чтобы уменьшить фотообесцвечивание флуоресцентного репортера, запись выполнялась с использованием 6-минутного рабочего цикла 83% (5 мин вкл/1 мин выкл). Базовый сигнал дофамина во время движения в открытом поле регистрировали в течение 20 мин. Затем мышам вводили ДЦЗ (100 мкг/кг, подкожно) и возвращали в камеру открытого поля для регистрации дофаминовой флуоресценции еще в течение 60 мин.
Данные фотометрии волокон были проанализированы с использованием пользовательского кода, написанного в RStudio (GitHub — xiaojieliu17/Fiber-photometry) (Snarrenberg, 2022). Чтобы рассчитать временной ряд ΔF/F, к данным была применена линейная аппроксимация, и подобранный сигнал 560 нм был выровнен и вычтен из сигнала 465 нм, а затем разделен на подобранный сигнал 560 нм, чтобы получить значения ΔF/F. Для анализа частоты дофаминовых событий обработанные значения ΔF/F были перенесены в файл ABF с использованием Minianalysis (Bluecell Co., Сеул, Корея). Затем частота переходных процессов дофамина была проанализирована с помощью их программного обеспечения Minianalysis. Начало события определяется 9Значение балла 0004 Z больше среднего плюс трехкратное стандартное отклонение баллов Z зарегистрированной флуоресценции в течение предшествующего периода 0,5 с, и результирующая пиковая частота сравнивалась между группами (Meng et al., 2018). После завершения экспериментов экспрессию AAV в VTA и NAc, а также локализацию имплантации оптического волокна в NAc проверяли с помощью иммуногистохимии.
Подготовка острых срезов головного мозга
Запросить подробный протокол
9Мышей 0002 WT и Epac2-cKO любого пола использовали для FSCV, электрофизиологических записей и scRT-PCR. Мышей анестезировали ингаляцией изофлурана и обезглавливали. Мозг помещали в агарозу с низкой температурой гелеобразования, а корональные полосатые срезы (толщиной 200 мкм) вырезали с помощью вибрирующего слайсера (Leica VT1200s, Nussloch, Германия), как описано в наших предыдущих исследованиях (Vickstrom et al., 2020). . Срезы готовили в растворе на основе холина, содержащем (в мМ): 92 NMDG, 2,5 KCl, 1,25 NaH 2 PO 4 , 0,5 CaCl 2 , 7 MgSO 4 , 26 NaHCO 3 , 25 глюкоза, 20 HEPES, 5 аскорбат натрия, 2 тиомочевина и 3 пируват натрия. Срезы немедленно переносили в искусственную спинномозговую жидкость (ИЦЖ) (в мМ): 119 NaCl, 2,5 KCl, 2,5 CaCl 2 , 1 MgCl 2 , 1,25 NaH 2 PO 4 9 3 9 32 NaHCO , и 10 глюкозы. Срезы давали восстановиться по крайней мере за 1 час до записи. Все растворы были насыщены 95% O 2 и 5% CO 2 .
Электрофизиология и scRT-PCR
Запросить подробный протокол
Для одновременного получения электрофизиологических данных и данных генетической экспрессии были выполнены записи прикрепленных к клеткам и целых клеток. Все оборудование очищали с помощью RNase Zap перед каждым использованием. Стеклянные пипетки прокаливали при 400°C не менее 4 часов для дезактивации РНКазы. Стеклянные пипетки (3–5 МОм) заполняли внутренним раствором, не содержащим РНКазы, содержащим (в мМ): 140 K-глюконата, 10 KCl, 10 HEPES, 0,2 EGTA, 2 MgCl 2 , 4 Mg-АТФ, 0,3 Na 2 GTP, 10 Na 2 -фосфокреатин (pH 7,2 с КОН). Внутренний раствор готовили на воде без РНКазы, делили на аликвоты и хранили при –80°С. Перед каждым экспериментом к 390 мкл внутреннего раствора добавляли 10 мкл рекомбинантного ингибитора РНКазы (1 ед/мкл, Sigma-Aldrich, Сент-Луис, Миссури) для увеличения выхода РНК. Записи целых клеток и прикрепленных к клеткам патч-зажимов были сделаны с использованием усилителей патч-зажим (Multiclamp 700B; Molecular Devices, Сан-Хосе, Калифорния) под инфракрасной дифференциально-интерференционной контрастной микроскопией. Сбор и анализ данных выполняли с использованием дигитайзеров DigiData 1440A и 1550B и программного обеспечения для анализа pClamp 10 (Molecular Devices). Сигналы фильтровались на частоте 2 кГц и дискретизировались на частоте 10 кГц. Активацию потенциала действия (ПД) регистрировали в прикрепленной к клетке конфигурации в присутствии CNQX (10 мкМ), D-AP5 (20 мкМ) и пикротоксина (50 мкМ) для блокирования возбуждающей и тормозной синаптической передачи, как описано (Vickstrom и др., 2021). Потенциалы соединения между накладной пипеткой и ванной ACSF аннулировали перед получением герметика. Емкость мембраны измеряли с помощью программного обеспечения Clampex (Molecular Devices) с использованием шагов гиперполяризации и деполяризации малой амплитуды (±5 мВ). Последовательное сопротивление (10–20 МОм) контролировалось на протяжении всех записей, и данные отбрасывались, если сопротивление изменялось более чем на 20%. Все записи были выполнены при 32 ± 1 ° C с использованием автоматического регулятора температуры (Warner Instruments LLC, Hamden, CT).
Содержимое клеток аспирировали в стеклянную пипетку, применяя слабое отрицательное давление в конце каждой записи. Во время процесса аспирации сохранялось уплотнение Gigaohm, чтобы уменьшить внешнее загрязнение. После аспирации пипетку медленно извлекали из ячейки. Затем содержимое клеток распределяли в микроцентрифужную пробирку без РНКазы при положительном давлении. Комплементарную ДНК (кДНК) синтезировали из цитоплазматической мРНК с использованием набора для синтеза кДНК iScript Advanced (Bio-Rad Laboratories, Hercules, CA). Затем гены-мишени амплифицировали с помощью двух стадий ПЦР с использованием следующих пар праймеров. Gapdh , прямой 5’-AGCTTGTCATCAACGGGAAG-3’, обратный 5’-GTCATGAGCCCTTCCACAAT-3’, 331 п. н.; Th , прямой 5’-TGCCAGAGAAGGACAAGGTTC-3’, обратный 5’-CGATACGCCTGGTCAGAGA-3’, 131 п.н.; Gad2 , прямой 5’-GTTCCTTTCCTGGTGAGTGC-3’, обратный 5’-TGCATCAGTCCCTCCTCTCTCT-3’, 266 п.н.; Epac2 , прямой 5’-TGGAACCAACTGGTATGCTG-3’, обратный 5’-CCAATTCCCAGAGTGCAGAT-3’, 102 п.н.; Epac1 , прямой 5’-GGACAAAGTCCCCTACGACA-3’, обратный 5’-CTTGGTCCAGTGGTCCTCAT-3’, 121 п.н.; Slc17a6 , прямой 5’-CGCTGCTTCTGGTTGTTGGC-3’, обратный 5’-AAACCATCCCCGACAGCGTG-3’, 186 п.н. Каждая ПЦР-амплификация состояла из 35 циклов: 94°С — 30 с, 58°С — 45 с, 72°С — 1 мин. Продукты второй ПЦР анализировали в 1,5% агарозных гелях с использованием бромистого этидия.
Циклическая вольтамперометрия с быстрым сканированием
Запросить подробный протокол
Микроэлектрод из цилиндрического углеродного волокна (диаметром 7 мкм, Гудфеллоу, Окдейл, Пенсильвания) с открытой конечной длиной 100–150 мкм был опущен в четыре разных участка стриатума, DMS, DLS, ядра NAc и оболочки NAc. в стриарных срезах. Координаты определяли путем визуального осмотра среза с помощью атласа мозга мыши (переднезадний, 0,9–1,7 мм). Микроэлектрод заполняли раствором, содержащим 150 мМ KCl. Треугольные сигналы (удерживающиеся при –0,4 В, от –0,4 до 1,3 В при 400 В/с) применялись каждые 100 мс с использованием программного обеспечения Demon (Yorgason et al., 2011) и потенциостата Chem-Clamp (Dagan Corporation, Миннеаполис, Миннесота). ). Стимулирующий электрод помещали на расстоянии ~100 мкм от микроэлектрода из углеродного волокна, и высвобождение дофамина вызывали одиночным импульсом электрического стимула (250 мкА, длительность 0,2 мс) или последовательностью стимулов из 5 импульсов с частотой 100 Гц (Liu et al., 2018b). ). После записи электроды калибровали в 0,1–2 мкМ дофамина в ACSF. Высвобождение дофамина вызывали одиночной импульсной стимуляцией (250 мкА, длительность 0,2 мс) каждые 2 мин, исследовали влияние агонистов и/или антагонистов Epac2 или PKA на высвобождение дофамина. После стабильной фоновой записи в течение 10 минут их влияние на вызванное высвобождение дофамина исследовали еще в течение 40 минут. Все записи проводились при 32 ± 1°С с использованием автоматического регулятора температуры (Warner Instruments LLC).
Иммунофлуоресцентное окрашивание
Запросить подробный протокол
Мышей анестезировали кетамином (90 мг/кг, внутрибрюшинно) и ксилазином (10 мг/кг, внутрибрюшинно) и транскардиально перфузировали 0,1 М раствором фосфата натрия, забуференным физиологическим раствором (PBS), а затем 4% параформальдегидом в 4% растворе сахарозы-PBS (pH 7.4). После перфузии мозг извлекали и дополнительно фиксировали в том же фиксаторе на 4 ч при 4°С и обезвоживали в возрастающих концентрациях сахарозы (20% и 30%) в 0,1 М PBS при 4°С. Фиксированный мозг замораживали на сухом льду и вырезали коронарные VTA или полосатые срезы (20 мкм) с помощью криостата Leica (CM1860). Срезы VTA инкубировали с первичными антителами против тирозингидроксилазы (TH, кроличьи поликлональные, 1:300, Santa Cruz Biotechnology, Inc, Dallas, TX) при 4°C в течение 48 часов. После трехкратной промывки PBS по 15 минут срезы VTA затем инкубировали во вторичном антителе: козьем антикроличьем IgG Alexa Fluor-488 в течение 4 часов при комнатной температуре в темноте. Корональные стриарные срезы были регидратированы в PBS и пермеабилизированы 0,1% Triton X-100 в PBS с последующей инкубацией в течение 20 мин с темно-красным флуоресцентным окрашиванием Ниссля 640/660 (N-21483). Конфокальное изображение было выполнено с использованием вертикального конфокального микроскопа Leica SP8.
Химические вещества
ESI-05, натриевая соль 6-Bnz-цАМФ, дигидрохлорид H-89 были приобретены у Tocris Bioscience (Миннеаполис, Миннесота). Sp-8-BnT-cAMPS (S-220) был приобретен в BioLog Life Science Institute (Фармингдейл, Нью-Йорк). Водорастворимый DCZ был приобретен у Hello Bio Inc (Принстон, Нью-Джерси). Кокаин HCl был предоставлен Программой снабжения лекарствами NIDA. Все другие распространенные химикаты были приобретены у Sigma-Aldrich.
Статистический анализ
Запросить подробный протокол
Данные представлены как среднее ± SEM. Наборы данных сравнивали либо с t -критерием Стьюдента, либо с парным t -тестом, либо с однофакторным дисперсионным анализом, либо с двусторонним дисперсионным анализом повторных измерений с последующим апостериорным анализом Тьюки. Для непараметрических данных контрольных точек PR было проведено сравнение с ранговым критерием Манна-Уитни или однофакторным дисперсионным анализом Крускала-Уоллиса для рангов с последующим апостериорным анализом Данна. Результаты считались значимыми при p<0,05.
Доступность данных
Все данные, полученные или проанализированные в ходе этого исследования, включены в рукопись и вспомогательный файл; Файлы исходных данных предоставлены для рисунков 1-7. Пользовательский код, используемый для анализа данных фотометрии волокна, доступен по адресу https://github.com/xiaojieliu17/Fiber-photometry (копия заархивирована по адресу swh:1:rev:ea959a0aed0ca49498c77d4d5479d004b67
).
Каталожные номера
- Акуна С
- Лю Х
- Зюдхоф ТК
(2016)
Как сделать активную зону: неожиданная универсальная функциональная избыточность между ободками и RIM-bps
Нейрон 91 :792–807.
https://doi.org/10.1016/j.neuron.2016.07.042
- пабмед
- Google Scholar
- Андерсон СМ
- Пирс RC
(2005)
Вызванные кокаином изменения в передаче сигналов дофаминовых рецепторов: последствия для подкрепления и восстановления
Фармакология и терапия 106 :389–403.
https://doi.org/10.1016/j.pharmthera.2004.12.004
- пабмед
- Google Scholar
- Андерсон EM
- Ларсон EB
- Гусман Д
- Виссман АМ
- Неве РЛ
- Нестлер EJ
- Сам ДВ
(2018)
Сверхэкспрессия гистон-диметилтрансферазы g9a в оболочке прилежащего ядра увеличивает самоуправление кокаином, вызванное стрессом восстановление и тревогу
The Journal of Neuroscience 38 :803–813.
https://doi.org/10.1523/JNEUROSCI.1657-17.2017
- пабмед
- Google Scholar
- Баамер F
- Сингхмар П
- Чжоу И
- Хэнкок JF
- Ченг Х
- Хейнен CJ
- Кавеларс А
(2016)
Epac1 взаимодействует с импортином β1 и контролирует рост нейритов независимо от цАМФ и rap1
Scientific Reports 6 :36370.
https://doi.org/10.1038/srep36370
- пабмед
- Google Scholar
- Баррьентос С
- Ноуленд D
- Ву MMJ
- Лилашароен V
- Хуан КВ
- Маленькая РЦ
- Лим БК
(2018)
Вызванная кокаином структурная пластичность во входных областях различных типов клеток в прилежащем ядре
Биологическая психиатрия 84 :893–904.
https://doi.org/10.1016/j.biopsych.2018.04.019
- пабмед
- Google Scholar
- Бенуа-Маранд М
- Джабер М
- Гонон F
(2000)
Высвобождение и элиминация дофамина in vivo у мышей, лишенных переносчика дофамина: функциональные последствия
The European Journal of Neuroscience 12 :2985–2992.
https://doi.org/10.1046/j.1460-9568.2000.00155.x
- пабмед
- Google Scholar
- Качопе R
- Матео Y
- Матур БН
- Ирвинг Дж
- Ван HL
- Моралес М
- Любовник ДМ
- Ура JF
(2012)
Избирательная активация холинергических интернейронов усиливает аккумулятивное фазовое высвобождение дофамина: задает тон для обработки вознаграждения
Отчеты сотовых 2 :33–41.
https://doi.org/10.1016/j.celrep.2012.05.011
- пабмед
- Google Scholar
- Кэхилл ME
- Багот RC
- Ганкарз АМ
- Уокер ДМ
- Солнце H
- Ван ZJ
- Хеллер EA
- Фэн Дж
- Кеннеди PJ
- Ку JW
- Кейтс HM
- Неве РЛ
- Шен Л
- Дитц DM
- Нестлер EJ
(2016)
Двунаправленная синаптическая структурная пластичность после хронического приема кокаина происходит за счет передачи сигналов малой гтпазы rap1
Нейрон 89 :566–582.
https://doi.org/10.1016/j.neuron.2016.01.031
- пабмед
- Google Scholar
- Кейн SB
- Негус СС
- Мелло НК
- Патель S
- Бристоу Л
- Кулаговский J
- Валлоне Д
- Сайарди А
- Боррелли E
(2002)
Роль дофаминовых D2-подобных рецепторов в самоуправлении кокаином: исследования с мутантными мышами рецептора D2 и новыми антагонистами рецептора D2
The Journal of Neuroscience 22 :2977–2988.
https://doi.org/10.1523/JNEUROSCI.22-07-02977.2002
- пабмед
- Google Scholar
- Карелли RM
(2002)
Активация клеток прилежащего ядра во время целенаправленного поведения при употреблении кокаина в сравнении с «естественным» подкреплением
Физиология и поведение 76 :379–387.
https://doi.org/10.1016/s0031-9384(02)00760-6
- пабмед
- Google Scholar
- Чен Р
- Макинтош S
- Хемби SE
- Солнце H
- Секстон Т
- Мартин TJ
- Чайлдерс SR
(2018)
Высокие и низкие дозы кокаина по-разному регулируются дофаминовыми рецепторами D2 в вентральной области покрышки и прилежащем ядре
Neuroscience Letters 671 :133–139.
https://doi.org/10.1016/j.neulet.2018.02.026
- пабмед
- Google Scholar
- Кристенсен AE
- Сельхейм F
- де Ройдж J
- Дремьер S
- Шведе F
- Дао КК
- Мартинес А
- Менхаут C
- Бос JL
- Генизер HG
- Доскеланд SO
(2003)
Аналоговое картирование CAMP epac1 и киназы CAMP: отличительные аналоги демонстрируют, что epac и киназа CAMP действуют синергетически, способствуя удлинению нейритов клеток PC-12
https://doi.
org/10.1074/jbc.M302179200- пабмед
- Google Scholar
- Корре J
- фургон Зессен R
- Лоурейро М
- Патриарх Т
- Тянь Л
- Пасколи В
- Люшер C
(2018)
Дофаминовые нейроны, проецирующиеся на медиальную оболочку прилежащего ядра, вызывают подкрепление героина
eLife 7 :e39945.
https://doi.org/10.7554/eLife.39945
- пабмед
- Google Scholar
- де Ройдж J
- Зварткруис FJ
- Верхейен MH
- Крутой РХ
- Нейман СМ
- Виттингхофер А
- Бос JL
(1998)
Epac представляет собой фактор обмена гуаниновых нуклеотидов rap1, непосредственно активируемый циклическим AMP 9.
0007Природа 396 :474–477.
https://doi.org/10.1038/24884
- пабмед
- Google Scholar
- Ди Кьяра G
- Императо А
(1988)
Наркотики, которыми злоупотребляют люди, преимущественно увеличивают концентрацию синаптического дофамина в мезолимбической системе свободно движущихся крыс
PNAS 85 :5274–5278.
https://doi.org/10.1073/pnas.85.14.5274
- пабмед
- Google Scholar
- Донг Ю
- Тейлор младший
- Волк ME
- Шахам Й
(2017)
Цепь и механизмы синаптической пластичности при рецидиве приема наркотиков
The Journal of Neuroscience 37 :10867–10876.
https://doi.org/10.1523/JNEUROSCI.1821-17.2017
- пабмед
- Google Scholar
- Фернандес ХБ
- Риордан S
- Номура Т
- Реммерс CL
- Краниотис S
- Маршалл JJ
- Кукрея Л
- Вассар Р
- Подрядчик А
(2015)
Epac2 опосредует цАМФ-зависимую потенциацию нейротрансмиссии в гиппокампе
Журнал неврологии 35 :6544–6553.
https://doi.
org/10.1523/JNEUROSCI.0314-14.2015- пабмед
- Google Scholar
- Ферреро JJ
- Альварес AM
- Рамирес-Франко Х
- Годино МС
- Бартоломе-Мартин D
- Агуадо C
- Торрес М
- Лухан Р
- Сируэла F
- Санчес-Прието J
(2013)
β-адренергические рецепторы активируют обменный белок, непосредственно активируемый цАМФ (epac), транслоцируют munc13-1 и усиливают взаимодействие rab3a-rim1α, потенцируя высвобождение глутамата в окончаниях цереброкортикальных нервов
Журнал биологической химии 288 : 31370–31385.
https://doi.org/10.1074/jbc.M113.463877
- пабмед
- Google Scholar
- Гайнетдинов Р.Р.
- Джонс SR
- Фумагалли Ф
- Вайтман RM
- Кэрон МГ
(1998)
Переоценка роли переносчика дофамина в гомеостазе дофаминовой системы
Brain Research. Brain Research Reviews 26 : 148–153.
https://doi.org/10.1016/s0165-0173(97)00063-5
- пабмед
- Google Scholar
- Гутьеррес-Кастельянос N
- Да Сильва-Матос CM
- Чжоу К
- Песнь CB
- Реннер МС
- Коене LMC
- Озйылдырым О
- Шпренгель R
- Кесселс HW
- Де Зеув CI
(2017)
Моторное обучение требует синаптической потенциации клеток Пуркинье посредством активации субъединицы АМРА-рецептора glua3
Нейрон 93 :409–424.
https://doi.org/10.1016/j.neuron.2016.11.046
- пабмед
- Google Scholar
- Газонокосилка RC
- Сала-Ньюби ГБ
- Ву YJ
- Ньюбай AC
- Бонд М
(2011)
PKA и epac синергетически ингибируют пролиферацию гладкомышечных клеток
Журнал молекулярной и клеточной кардиологии 50 : 87–98.
https://doi.org/10.1016/j.yjmcc.2010.10.010
- пабмед
- Google Scholar
- Хиранита Т
(2015)
Открытие лекарств: важность оценки кривых доза-эффект при самостоятельном введении лекарств
Журнал алкоголизма и наркотической зависимости 3 :e121.
https://doi.org/10.4172/2329-6488.1000e121
- пабмед
- Google Scholar
- Каливас PW
(2018)
Деконструкция и реконструкция дихотомии нейронов, экспрессирующих дофаминовый рецептор-1 и дофаминовый рецептор-2
Биологическая психиатрия 84 :862–864.
https://doi.org/10.1016/j.biopsych.2018.10.007
- пабмед
- Google Scholar
- Кавасаки Н
- Спрингетт GM
- Мочизуки N
- Токи С
- Накая М
- Мацуда М
- Хаусман DE
- Грейбиэль AM
(1998)
Семейство цАМФ-связывающих белков, которые непосредственно активируют rap1
Science 282 :2275–2279.
https://doi.org/10.1126/science.282.5397,2275
- пабмед
- Google Scholar
- Кек ТМ
- Цзоу МФ
- Би GH
- Чжан HY
- Ван XF
- Ян ДЖ
- Шривастава Р
- Гарднер EL
- Си ZX
- Ньюман AH
(2014)
Новый антагонист mglur5, MFZ 10-7, ингибирует поведение, связанное с приемом кокаина и поиском кокаина у крыс
Addiction Biology 19 :195–209.
https://doi.org/10.1111/adb.12086
- пабмед
- Google Scholar
- Кооб ГФ
- Блум FE
(1988)
Клеточные и молекулярные механизмы лекарственной зависимости
Наука 242 :715–723.
https://doi.org/10.1126/science.20
- пабмед
- Google Scholar
- Лоран AC
- Бреклер М
- Бертхаус М
- Лезуальк F
(2012)
Роль epac в мозге и сердце
Biochemical Society Transactions 40 :51–57.
https://doi.org/10.1042/BST20110642
- пабмед
- Google Scholar
- Леви Д
- Шабат-Симон М
- Шалев У
- Барнеа-Игаэль N
- Купер А
- Занген А
(2007)
Повторная электрическая стимуляция областей мозга, связанных с вознаграждением, воздействует на кокаин, но не на «естественное» подкрепление
The Journal of Neuroscience 27 :14179–14189.
https://doi.org/10.1523/JNEUROSCI.4477-07.2007
- пабмед
- Google Scholar
- Лю Икс
- Чен Ю
- Тонг Дж
- Рейнольдс AM
- Праудфут SC
- Ци Дж
- Пензес П
- Лу Ю
- Лю QS
(2016)
Передача сигналов Epac необходима для индуцированного кокаином изменения состава субъединиц рецептора AMPA в вентральной области покрышки
The Journal of Neuroscience 36 :4802–4815.
https://doi.org/10.1523/JNEUROSCI.3186-15.2016
- пабмед
- Google Scholar
- Лю С
- Кершберг Л
- Ван Дж
- Шнеебергер S
- Кезер ПС
(2018а)
Секреция дофамина опосредована редкими активными зонами, подобными сайтам высвобождения
Сотовый 172 :706–718.
https://doi.org/10.1016/j.cell.2018.01.008
- пабмед
- Google Scholar
- Лю Икс
- Ли Ю
- Ю Л
- Викстром КР
- Лю QS
(2018б)
Передача сигналов VTA mtor регулирует динамику дофамина, кокаин-индуцированные синаптические изменения и вознаграждение
https://doi.
org/10.1038/npp.2017.247- пабмед
- Google Scholar
- Лю С
- Кезер ПС
(2019)
Механизмы и регуляция высвобождения дофамина
Современное мнение в области нейробиологии 57 : 46–53.
https://doi.org/10.1016/j.conb.2019.01.001
- пабмед
- Google Scholar
- Лю Икс
- Ю Х
- Чен Б
- Фридман В
- Му Л
- Келли TJ
- Руис-Перес G
- Чжао Л
- Бай Х
- Хиллард CJ
- Лю QS
(2022)
Агонист CB2 gw842166x защищает от 6-OHDA-индуцированного анксиогенного и депрессивного поведения у мышей
Biomedicines 10 :1776.
https://doi.org/10.3390/биомедицина10081776
- пабмед
- Google Scholar
Препринт
- Малер СВ
- Бродник ЗД
- Кокс БМ
- Бухта унитаз
- Бенцли БС
- Коуп ZA
- Лин ЕС
- Риди MD
- Скофилд, доктор медицины
- Мессингер J
- Ригель AC
- Испания RA
- Астон-Джонс G
(2018)
Хемогенетические манипуляции с дофаминовыми нейронами вентральной сегментарной области раскрывают многогранную роль в злоупотреблении кокаином
биоRxiv.
https://doi.org/10.1101/246595
- Google Scholar
- Мартин Р
- Гарсия-Фонт N
- Суарес-Пинилья AS
- Бартоломе-Мартин D
- Ферреро JJ
- Лухан Р
- Торрес М
- Санчес-Прието J
(2020)
β-адренергические рецепторы/передача сигналов epac увеличивают размер легко высвобождаемого пула синаптических пузырьков, необходимых для LTP параллельных волокон
The Journal of Neuroscience 40 :8604–8617.
https://doi.
org/10.1523/JNEUROSCI.0716-20.2020- пабмед
- Google Scholar
- Мелло НК
- Негус СС
(1996)
Доклиническая оценка фармакотерапии для лечения злоупотребления кокаином и опиоидами с использованием процедур самостоятельного приема наркотиков
Нейропсихофармакология 14 :375–424.
https://doi.org/10.1016/0893-133X(95)00274-H
- пабмед
- Google Scholar
- Мэн С
- Чжоу Дж
- Папанери А
- Педдада Т
- Сюй К
- Цуй Г
(2018)
Фотометрия волокон со спектральным разрешением для многокомпонентного анализа мозговых цепей
Нейрон 98 :707–717.
https://doi.org/10.1016/j.neuron.2018.04.012
- пабмед
- Google Scholar
- Мереу М
- Хиранита Т
- Иордания CJ
- Чун LE
- Лопес JP
- Коджиано MA
- Квартирник JC
- Би GH
- Кейрон JD
- Си ZX
- Ньюман AH
- Кац JL
- Танда G
(2020)
Модафинил усиливает самостоятельный прием кокаина за счет дофамин-независимого механизма: возможное вовлечение щелевых контактов
Нейропсихофармакология 45 :1518–1526.
https://doi.org/10.1038/s41386-020-0680-5
- пабмед
- Google Scholar
- Му Л
- Лю Икс
- Ю Х
- Ху М
- Фридман В
- Келли TJ
- Чжао Л
- Лю QS
(2021)
Ибудиласт ослабляет самостоятельный прием кокаина и восстановление поиска кокаина у крыс, вызванное стимулом и сигналом
.
https://doi.org/10.1016/j.neuropharm.2021.108830
- пабмед
- Google Scholar
- Нагаи Y
- Миякава N
- Такува Х
- Хори Ю
- Ояма К
- Джи Б
- Такахаши М
- Хуан XP
- Слокум СТ
- ДиБерто JF
- Сюн Ю
- Урушихата Т
- Хирабаяси Т
- Фудзимото А
- Мимура К
- Английский JG
- Лю Дж
- Иноуэ К-И
- Кумата К
- Секи С
- Оно М
- Симодзё М
- Чжан М-Р
- Томита Y
- Накахара Дж
- Сухара Т
- Такада М
- Хигучи М
- Джин Дж
- Рот БЛ
- Минамимото Т
(2020)
Дехлорклозапин, мощный и селективный хемогенетический активатор, обеспечивает быструю нейронную и поведенческую модуляцию у мышей и обезьян
Nature Neuroscience 23 :1157–1167.
https://doi.org/10.1038/s41593-020-0661-3
- пабмед
- Google Scholar
- Нестлер EJ
- Тервиллигер РЗ
- Уокер младший
- Севарино КА
- Думан РС
(1990)
Хроническое лечение кокаином снижает уровни субъединиц G-белка gi-альфа и го-альфа в отдельных областях мозга крыс
Journal of Neurochemistry 55 :1079–1082.
https://doi.org/10.1111/j.1471-4159.1990.tb04602.x
- пабмед
- Google Scholar
- Нестлер EJ
(2001)
Молекулярная основа долговременной пластичности, лежащей в основе зависимости
Nature Reviews.
Неврология 2 : 119–128.https://doi.org/10.1038/35053570
- пабмед
- Google Scholar
- Нестлер EJ
(2016)
Размышления о: «Общая роль адаптации G-белков и циклической системы AMP в опосредовании хронического действия морфина и кокаина на функцию нейронов».
Исследования мозга 1645 :71–74.
https://doi.org/10.1016/j.brainres.2015.12.039
- пабмед
- Google Scholar
- О’Коннор ЕС
- Чепмен К
- Батлер П
- Мед АН
(2011)
Прогностическая валидность модели самоконтроля крыс в отношении склонности к жестокому обращению
Нейронаука и биоповеденческие обзоры 35 :912–938.
https://doi.org/10.1016/j.neubiorev.2010.10.012
- пабмед
- Google Scholar
- Огава СК
- Ватабэ Учида М
(2018)
Организация дофаминовой и серотониновой системы: анатомо-функциональное картирование моносинаптических входов с использованием вируса бешенства
Фармакология, биохимия и поведение 174 :9–22.
https://doi.org/10.1016/j.pbb.2017.05.001
- пабмед
- Google Scholar
- Одзаки N
- Шибасаки Т
- Кашима Y
- Мики Т
- Такахаши К
- Уэно H
- Сунага Y
- Яно Х
- Мацуура Y
- Иванага Т
- Такай Y
- Сейно С
(2000)
CAMP-GEFII является прямой мишенью для цАМФ при регулируемом экзоцитозе
Nature Cell Biology 2 :805–811.
https://doi.org/10.1038/35041046
- пабмед
- Google Scholar
- Патриарх Т
- Чо JR
- Мертен К
- Хоу МВт
- Марли А
- Сюн WH
- Народный RW
- Бруссард GJ
- Лян Р
- Джанг MJ
- Чжун Х
- Домбек Д
- фон Застров M
- Ниммерьян А
- Градинару В
- Уильямс JT
- Тянь Л
(2018)
Сверхбыстрая нейронная визуализация динамики дофамина с помощью разработанных генетически кодируемых сенсоров
Наука 360 :eaat4422.
https://doi.org/10.1126/science.aat4422
- пабмед
- Google Scholar
- Перейра Л
- Ченг Х
- Лаосский DH
- На Л
- фургон Оорт RJ
- Коричневый JH
- Веренс XHT
- Чен Дж
- Берс ДМ
(2013)
Epac2 опосредует сердечную β1-адренергическую утечку Са2+ из саркоплазматического ретикулума и аритмию
Кровообращение 127 :913–922.
https://doi.
org/10.1161/CIRCULATIONAHA.12.148619- пабмед
- Google Scholar
- Ричардсон NR
- Робертс DC
(1996)
Схемы прогрессивного соотношения в исследованиях самостоятельного приема лекарств на крысах: метод оценки эффективности подкрепления
Journal of Neuroscience Methods 66 :1–11.
https://doi.org/10.1016/0165-0270(95)00153-0
- пабмед
- Google Scholar
- Робертс DCS
- Габриэле А
- Циммер БА
(2013)
Объединение реакций поиска кокаина и приема кокаина в экспериментах по внутривенному самоуправлению у крыс: методологические и интерпретационные соображения
Неврологические и биоповеденческие обзоры 37 :2026–2036.
https://doi.org/10.1016/j.neubiorev.2013.04.017
- пабмед
- Google Scholar
- Робертс БМ
- Дойг НМ
- Бримблкомб KR
- Лопес EF
- Сиддорн RE
- Трелфелл S
- Коннор-Робсон N
- Бенгоа-Верньори N
- Пастернак N
- Уэйд-Мартинс R
- Магилл PJ
- Крэгг SJ
(2020)
Транспортеры поглощения ГАМК поддерживают высвобождение дофамина в дорсальном полосатом теле с неадекватным подавлением в модели паркинсонизма
Nature Communications 11 :4958.
https://doi.org/10.1038/s41467-020-18247-5
- пабмед
- Google Scholar
- Робишо WG
- Ченг Х
(2018)
Внутриклеточный цАМФ-сенсор EPAC: физиология, патофизиология и развитие терапии
Physiological Reviews 98 :919–1053.
https://doi.org/10.1152/physrev.00025.2017
- пабмед
- Google Scholar
- Ромье P
- Хост Л
- Гобай S
- Санднер Г
- Аунис Д
- Цвиллер J
(2008)
Ингибиторы гистондеацетилазы снижают самостоятельный прием кокаина, но не сахарозы у крыс
The Journal of Neuroscience 28 :9342–9348.
https://doi.org/10.1523/JNEUROSCI.0379-08.2008
- пабмед
- Google Scholar
- Рот БЛ
(2016)
DREADD для неврологов
Нейрон 89 :683–694.
https://doi.org/10.1016/j.neuron.2016.01.040
- пабмед
- Google Scholar
- Шмидт HD
- Пирс RC
(2010)
Вызванная кокаином нейроадаптация при передаче глутамата: потенциальные терапевтические мишени для тяги и зависимости
Анналы Нью-Йоркской академии наук 1187 : 35–75.
https://doi.org/10.1111/j.1749-6632.2009.05144.х
- пабмед
- Google Scholar
- Шведе F
- Бертинетти D
- Лангерийс CN
- Хаддерс MA
- Венк H
- Элленбрук JH
- де Конинг EJP
- Бос JL
- Герберг FW
- Генизер H-G
- Янссен РАЙ
- Реманн H
(2015)
Структурно-ориентированный дизайн селективных агонистов epac1 и epac2
PLOS Биология 13 :e1002038.
https://doi.org/10.1371/journal.pbio.1002038
- пабмед
- Google Scholar
- Сам ДВ
(2004)
Регуляция поведения, связанного с приемом и поиском наркотиков, посредством нейроадаптаций в мезолимбической дофаминовой системе
Нейрофармакология 47 Приложение 1 : 242–255.
https://doi.org/10.1016/j.neuropharm.2004.07.005
- пабмед
- Google Scholar
- Шибасаки Т
- Сунага Y
- Фудзимото К
- Кашима Y
- Сейно С
(2004)
Взаимодействие АТФ-сенсора, цАМФ-сенсора, ca2+-сенсора и потенциалзависимого ca2+-канала при экзоцитозе инсулиновых гранул
The Journal of Biological Chemistry 279 :7956–7961.
https://doi.org/10.1074/jbc.M30
- пабмед
- Google Scholar
- Слоски ЛМ
- Бай Ю
- Тот К
- Рэй С
- Рошель ЛК
- Бадеа А
- Чандрасекар Р
- Погорелов В.М.
- Авраам DM
- Атлури N
- Педдибхотла С
- Хедрик MP
- Хершбергер П
- Мэлони П
- Юань H
- Ли З
- Ветсель WC
- Пинкертон AB
- Барак ЛС
- Кэрон МГ
(2020)
Связанный с β-аррестином аллостерический модулятор ntsr1 избирательно ослабляет аддиктивное поведение
Клетка 181 :1364–1379.
https://doi.org/10.1016/j.cell.2020.04.053
- Google Scholar
Препринт
- Слоски ЛМ
- Пирес А
- Бай Ю
- Кларк N
- Хаузер ER
- Валовой JD
- Свинина F
- Чжоу И
- Чен Х
- Погорелов В.М.
- Тот К
- Ветсель WC
- Барак ЛС
- Кэрон МГ
(2021)
Создание парадигмы многоэтапного внутривенного самоуправления у мышей
биоRxiv.
https://doi.org/10.1101/2020.11.25.398503
- Google Scholar
Программное обеспечение
- Снарренберг ST
(2022)
Волоконно-фотометрия, версия v.1.3.3.R
Github.
https://github.com/xiaojieliu17/Fiber-photometry
- Стриплин CD
- Каливас PW
(1992)
Корреляция между поведенческой сенсибилизацией к кокаину и АДФ-рибозилированием G-белка в вентральной области покрышки
Brain Research 579 :181–186.
https://doi.org/10.1016/0006-8993(92)-f
- пабмед
- Google Scholar
- Солнце F
- Чжоу Дж
- Дай Б
- Цянь Т
- Цзэн Дж
- Ли Х
- Чжо Ю
- Чжан И
- Ван И
- Цянь C
- Тан К
- Фэн Дж
- Донг Х
- Лин Д
- Цуй Г
- Ли Ю
(2020)
Сенсоры GRAB следующего поколения для мониторинга дофаминергической активности in vivo
Природные методы 17 :1156–1166.
https://doi.org/10.1038/s41592-020-00981-9
- пабмед
- Google Scholar
- Тенгхольм А
- Гюльфе E
(2017)
Передача сигналов CAMP при секреции инсулина и глюкагона
Диабет, ожирение и метаболизм 19 Приложение 1 :42–53.
https://doi.org/10.1111/dom.12993
- пабмед
- Google Scholar
- Трелфелл S
- Лалик Т
- Платт Нью-Джерси
- Дженнингс К. А.
- Дейссерот К
- Крэгг SJ
(2012)
Высвобождение дофамина в полосатом теле запускается синхронизированной активностью холинергических интернейронов
Нейрон 75 :58–64.
https://doi.org/10.1016/j.neuron.2012.04.038
- пабмед
- Google Scholar
- Тонг Дж
- Лю Икс
- Викстром С
- Ли Ю
- Ю Л
- Лу Ю
- Смрцкая АВ
- Лю QS
(2017)
Путь epac-фосфолипазы cε регулирует эндоканнабиноидную сигнализацию и индуцированное кокаином растормаживание дофаминовых нейронов вентральной области покрышки
The Journal of Neuroscience 37 :3030–3044.
https://doi.org/10.1523/JNEUROSCI.2810-16.2017
- пабмед
- Google Scholar
- Тритч NX
- Сабатини BL
(2012)
Дофаминергическая модуляция синаптической передачи в коре и стриатуме
Нейрон 76 :33–50.
https://doi.org/10.1016/j.neuron.2012.09.023
- пабмед
- Google Scholar
- Цалкова Т
- Мэй ФК
- Ли С
- Чепурный О. Г.
- Пиявка CA
- Лю Т
- Хольц ГГ
- Вудс ВЛ
- Ченг Х
(2012)
Изоформоспецифические антагонисты обменных белков, непосредственно активируемые цАМФ
PNAS 109 :18613–18618.
https://doi.org/10.1073/pnas.1210209109
- пабмед
- Google Scholar
- Викстром CR
- Лю Икс
- Чжан И
- Му Л
- Келли TJ
- Ян Х
- Ху ММ
- Снарренберг ST
- Лю QS
(2020)
Кальциевые каналы Т-типа способствуют взрывному возбуждению в субпопуляции нейронов медиальных крючков
ENeuro 7 :ENEURO.
0201-20.2020.https://doi.org/10.1523/ENEURO.0201-20.2020
- пабмед
- Google Scholar
- Викстром CR
- Лю Икс
- Лю С
- Ху ММ
- Му Л
- Ху Y
- Ю Х
- Любовь SL
- Хиллард CJ
- Лю QS
(2021)
Роль эндоканнабиноидной сигнализации в септогабенулярном пути в регуляции тревожного и депрессивного поведения
Молекулярная психиатрия 26 :3178–3191.
https://doi.org/10.1038/s41380-020-00905-1
- пабмед
- Google Scholar
- Волков НД
- Ван ГДж
- Теланг F
- Фаулер JS
- Логан Дж
- Чайлдресс AR
- Джейн М
- Ма Y
- Вонг С
(2006)
Сигналы кокаина и дофамин в дорсальном полосатом теле: механизм тяги при кокаиновой зависимости
The Journal of Neuroscience 26 :6583–6588.
https://doi.
org/10.1523/JNEUROSCI.1544-06.2006- пабмед
- Google Scholar
- Волков НД
- Ван ГДж
- Фаулер JS
- Томази Д
- Пресс-подборщик R
(2012)
Вознаграждение за еду и лекарства: перекрывающиеся цепи человеческого ожирения и зависимости
Текущие темы в поведенческой неврологии 11 : 1–24.
https://doi.org/10.1007/7854_2011_169
- пабмед
- Google Scholar
- Ван Дж
- Исикава М
- Ян Y
- Отака М
- Ким JY
- Гарднер GR
- Стефаник МТ
- Милованович М
- Хуан YH
- Ад JW
- Волк ME
- Шлютер ОМ
- Донг Ю
(2018)
Каскады гомеостатической дисрегуляции способствуют инкубации тяги к кокаину
Журнал неврологии 38 :4316–4328.
https://doi.org/10.1523/JNEUROSCI.3291-17.2018
- пабмед
- Google Scholar
- Волк ME
(2016)
Синаптические механизмы, лежащие в основе стойкой тяги к кокаину
Обзоры природы. Неврология 17 : 351–365.
https://doi.org/10.1038/nrn.2016.39
- пабмед
- Google Scholar
- Вулфри КМ
- Шривастава ДП
- Фотовала H
- Ямасита М
- Барболина М. В.
- Кэхилл ME
- Се Z
- Джонс К.А.
- Куиллиам Л.А.
- Пракрия М
- Пензес П
(2009)
Epac2 индуцирует ремоделирование синапсов и депрессию, а его формы, связанные с заболеванием, изменяют шипы
Nature Neuroscience 12 :1275–1284.
https://doi.org/10.1038/nn.2386
- пабмед
- Google Scholar
- Си ZX
- Песня R
- Ли Х
- Лу GY
- Пэн XQ
- Он Y
- Би GH
- Шэн СП
- Ян HJ
- Чжан Х
- Ли Дж
- Фроймовиц М
- Гарднер EL
(2017)
CTDP-32476: Перспективная агонистическая терапия для лечения кокаиновой зависимости
Нейропсихофармакология 42 :682–694.
https://doi.org/10.1038/npp.2016.155
- пабмед
- Google Scholar
- Ян Y
- Шу Х
- Лю Д
- Шан Ю
- Ву Ю
- Пей Л
- Сюй Х
- Тянь Q
- Чжан Дж
- Цянь К
- Ван YX
- Петралия RS
- Ту Вт
- Чжу LQ
- Ван JZ
- Лу Ю
(2012)
Нулевая мутация EPAC нарушает обучение и социальные взаимодействия посредством аномальной регуляции трансляции mir-124 и zif268
Нейрон 73 :774–788.
https://doi.org/10.1016/j.neuron.2012.02.003
- пабмед
- Google Scholar
- Йоргасон JT
- Испания RA
- Джонс SR
(2011)
Программное обеспечение для вольтамперометрии и анализа Demon: анализ вызванных кокаином изменений в передаче сигналов дофамина с использованием нескольких кинетических показателей
Journal of Neuroscience Methods 202 :158–164.
https://doi.org/10.1016/j.jneumeth.2011.03.001
- пабмед
- Google Scholar
- Циммер БА
- Добрин CV
- Робертс DCS
(2011)
Концентрация кокаина в мозге определяет дозу, которую крысы вводят себе самостоятельно по новой поведенческой схеме дозирования
Нейропсихофармакология 36 :2741–2749.
https://doi.org/10.1038/npp.2011.165
- пабмед
- Google Scholar
- Цинсмайер АК
- Донг Ю
- Хуан YH
(2022)
Кокаин-индуцированные проекционно-специфические и специфичные для типов клеток адаптации в прилежащем ядре
Молекулярная психиатрия 27 :669–686.
https://doi.org/10.1038/s41380-021-01112-2
- пабмед
- Google Scholar
00
Письмо-решение
Письмо с решением после рецензирования:
Благодарим вас за представление вашей статьи «Epac2 в дофаминовых нейронах среднего мозга способствует усилению кокаиновой зависимости за счет облегчения высвобождения дофамина» на рассмотрение eLife . Ваша статья была рассмотрена тремя рецензентами, а за оценкой наблюдали редактор-рецензент и Кейт Вассум в качестве старшего редактора. Следующее лицо, участвовавшее в рассмотрении вашей заявки, согласилось раскрыть свою личность: Эрин Калипари (Рецензент №1).
Рецензенты обсудили свои рецензии друг с другом, и редактор-рецензент составил черновой вариант, чтобы помочь вам подготовить исправленную заявку.
Существенные изменения:
Как указано ниже, рецензенты очень положительно оценили строгость и потенциальную значимость этой рукописи и высоко оценили включение множества различных технических подходов для определения роли пресинаптического Epac2 в передаче сигналов дофамина и подкреплении/мотивации кокаином. Несмотря на то, что критический анализ рецензента предлагал потенциальные эксперименты, которые могли бы помочь расширить и прояснить механистический вклад Epac2, общий консенсус заключался в том, что эти эксперименты не являются существенными для дальнейшего рассмотрения в eLife . Однако, как минимум, пересмотренная рукопись должна включать:
1) Разъяснение экспериментальных вопросов и опасений, поднятых каждым рецензентом, а также включение предлагаемых изменений текста.
2) Дополнительное или более подробное обсуждение ограничений, поднятых рецензентами, и последствия работы, включая обсуждение потенциально неспецифических эффектов манипуляций с Epac2 в NAc (Rev. 1), обоснование сосредоточения внимания на латеральной оболочке NAc , и обсуждение различий во вкладе Epac2 в вознаграждение сахарозой и кокаином (Rev. 2), и уточнение рабочей модели, представленной на рисунке 9.(Откр. 3).
Рецензент №1 (рекомендации для авторов):
1. Меня смущает, что в графике подкрепления FR1 вливания и жимы рычагами не совпадают. Если они отличаются, животное на самом деле не входит в схему FR1, поскольку каждая реакция на самом деле не приводит к вливанию препарата. Я бы просто изменил это на график с низкими усилиями или что-то в этом роде — никаких новых экспериментов не нужно было бы проводить.
2. Хотя эксперименты на рис. 6 интересны, важно обсудить, что это означает в контексте работы микросхемы. Возможно, что эффекты Epac2 могут избирательно оказывать локальное воздействие на дофаминовые окончания, независимо от соматических эффектов; однако дизайн не позволяет различать эффекты на дофаминовые окончания и другие системы нейротрансмиттеров в NAc, такие как ГАМК, ацетилхолин, глутамат и пептидергическое высвобождение. Если вы нокаутируете Epac2 в дофаминовых нейронах, вы по-прежнему ощущаете воздействие лекарств на локальные срезы? Рукопись содержит много данных, поэтому нет необходимости проводить эти дополнительные эксперименты, но это должно быть как минимум подробно обсуждено в обсуждении.
Рецензент №2 (рекомендации для авторов):
Рисунки 1 и 3: внутрибрюшинное введение ESI-05 ингибирует глобальный Epsac2. Учитывая обширные данные, основанные на манипуляциях с дофаминовыми нейронами, данные на этих рисунках не имеют существенного веса. Возможно, будет лучше, если некоторые результаты будут перемещены в раздел Suppl.
Рисунки 3 и 4: Интересно и несколько удивительно, что манипуляции с Epac2 влияли только на самостоятельный прием кокаина, не влияя на реакции, мотивированные сахарозой, учитывая, что дофамин также необходим для кондиционирования сахарозы. Любые идеи требуют обсуждения.
Рисунок 5: Боковая оболочка NAc была сфокусирована. Функционально медиальная оболочка преимущественно участвует в мотивированном поведении. Некоторое обоснование сосредоточения внимания на боковой оболочке поможет.
Рисунки 7 и 8: Возможно, следует рассмотреть контроли дикого типа. В частности, помимо мышей Epac2-cKO, будет ли хемогенетическое подавление/усиление высвобождения дофамина также влиять на мышей без генетических модификаций?
Рецензент №3 (Рекомендации авторам):
1. Чтобы ответить на вопрос № 1 в Public Review, потенциальный эксперимент, который авторы могут рассмотреть, состоит в том, чтобы вернуть белок Epac2 дикого типа в мутантные нейроны VTA DA (например, посредством сверхэкспрессии вирусного гена) с последующим измерением высвобождения DA. и кокаин С.А. Авторы также могут рассмотреть возможность тестирования, способствует ли хемогенная активация дофаминовых нейронов сахарозной СА, что может дать представление о природе и значении спасения. Если некоторые или все эти эксперименты неосуществимы или нецелесообразны, необходимо провести подробное обсуждение и/или обоснование для разъяснения.
2. Для решения пункта № 2 в Public Review авторы могут рассмотреть возможность повторения экспериментов по хемогенетическому ингибированию на мышах Epac2 cKO и посмотреть, блокирует ли делеция Epack2 или притупляет эффекты ингибирования высвобождения DA на кокаин SA у мутантных мышей, или предоставить больше подробные обсуждения или разъяснения.
3. Для решения пункта № 3 в публичном обзоре авторы должны представить временную динамику переходных изменений DA после введения DCZ и показать более репрезентативные записи сигнала DA.
4. Рабочая модель рисунка 9 может быть пересмотрена путем добавления некоторых промежуточных шагов между входящими пиками потенциала действия и стыковкой КА.
https://doi.org/10.7554/eLife.80747.sa1
Ответ автора
Рецензент №1 (Рекомендации авторам):
1. Меня смущает то, что в графике подкрепления FR1 вливания и жим рычага не совпадают. Если они отличаются, животное на самом деле не входит в схему FR1, поскольку каждая реакция на самом деле не приводит к вливанию препарата. Я бы просто изменил это на график с низкими усилиями или что-то в этом роде — никаких новых экспериментов не нужно было бы проводить.
Расписание FR1 имеет короткий период ожидания (10 секунд) после каждого активного (вознаграждаемого) тыкания в нос, во время которого тыкание в нос не приводит к вливанию кокаина. В результате количество уколов носом несколько превышает количество вливаний кокаина. Мы разъяснили это в условных обозначениях к рис. 1. Однако, чтобы сохранить соглашение, мы не изменили «график FR1» на «график с низкими усилиями».
2. Хотя эксперименты на рис. 6 интересны, важно обсудить, что это означает в контексте работы микросхемы. Возможно, что эффекты Epac2 могут избирательно оказывать локальное воздействие на дофаминовые окончания, независимо от соматических эффектов; однако дизайн не позволяет различать эффекты на дофаминовые окончания и другие системы нейротрансмиттеров в NAc, такие как ГАМК, ацетилхолин, глутамат и пептидергическое высвобождение. Если вы нокаутируете Epac2 в дофаминовых нейронах, вы по-прежнему ощущаете воздействие лекарств на локальные срезы? Рукопись содержит много данных, поэтому нет необходимости проводить эти дополнительные эксперименты, но это должно быть как минимум подробно обсуждено в обсуждении.
Мы согласны с рецензентом в том, что агонист Epac2 действует локально на окончания дофаминовых аксонов, поскольку разрезание среза удаляет сому дофаминовых нейронов. Используемый здесь подход Cre-LoxP приводил к полному нокауту Epac2 из целых дофаминовых нейронов среднего мозга (сомы и аксональные окончания). Важно отметить, что этот специфический нокаут дофаминовых нейронов полностью отменяет эффект агониста Epac2 на увеличение высвобождения дофамина, что позволяет предположить, что его эффекты не опосредованы другими системами нейротрансмиттеров. Мы добавили следующее обсуждение в исправленную рукопись (страницы 16-17, строки 372-381), чтобы еще больше прояснить и рассмотреть этот момент.
«Существует вероятность того, что агонист Epac2 S-220 повлиял на другие системы нейротрансмиттеров, косвенно модулируя высвобождение дофамина. Оптогенетическая стимуляция холинергических интернейронов в NAc индуцировала высвобождение дофамина посредством активации никотиновых ацетилхолиновых рецепторов (nAChR) (Cachope et al., 2012; Threlfell et al., 2012). ГАМК тонически подавляет высвобождение дофамина аксонами в дорсальном полосатом теле через ГАМК- и ГАМК-рецепторы (Roberts et al., 2020). Таким образом, ацетилхолин и ГАМК оказывают косвенное, но существенное влияние на выброс дофамина. Однако S-220-индуцированное усиление высвобождения дофамина устранялось Epac2-cKO из дофаминовых нейронов, что позволяет предположить, что дофаминовые окончания аксонов являются преобладающим местом действия S-220. В будущих исследованиях было бы интересно проверить, изменяется ли S-220-индуцированное усиление высвобождения дофамина блокаторами nAChR и рецепторов GABA».
Рецензент №2 (Рекомендации авторам):
Рисунки 1 и 3: внутрибрюшинное введение ESI-05 ингибирует глобальный Epsac2. Учитывая обширные данные, основанные на манипуляциях с дофаминовыми нейронами, данные на этих рисунках не имеют существенного веса. Возможно, будет лучше, если некоторые результаты будут перемещены в раздел Suppl.
Мы последовали совету рецензента и переместили рисунок 3 (ESI-05 о самостоятельном введении сахарозы) в дополнительный раздел (теперь рисунок 3 — приложение к рисунку 1). Мы хотели бы сохранить исходный рисунок 1, поскольку считаем, что ингибирование ESI-05 самостоятельного приема кокаина является основным открытием рукописи.
Рисунки 3 и 4: Интересно и несколько удивительно, что манипуляции с Epac2 влияли только на самостоятельный прием кокаина, не влияя на реакции, мотивированные сахарозой, учитывая, что дофамин также необходим для кондиционирования сахарозой.
Мы добавили следующее обсуждение в исправленную рукопись (страницы 15-16, строки 346-358):
«Кокаин и сахароза повышают уровень дофамина, уменьшая его поглощение и увеличивая его высвобождение, соответственно (Anderson and Pierce, 2005; Патриархи и др., 2018). Было несколько неожиданно, что ESI-05 и Epac2 ослабляли самостоятельный прием кокаина, но существенно не изменяли самостоятельный прием сахарозы. Однако сообщалось об аналогичной дифференциальной модуляции самостоятельного приема кокаина и сахарозы фармакологическими агентами (Levy et al., 2007; Romieu et al., 2008; Mu et al., 2021). Хотя в нейронных цепях, кодирующих вознаграждение за еду и наркотики, существует значительное перекрытие (Volkow et al., 2012), большинство нейронов NAc демонстрировали разные паттерны возбуждения в ответ на вознаграждение за еду и кокаин (Carelli, 2002). Как упоминалось во введении, повторное употребление кокаина приводит к снижению G 9экспрессия 1293 αi/o в VTA и активация передачи сигналов цАМФ (Nestler et al. , 1990; Striplin and Kalivas, 1992). Вполне вероятно, что Epac2 предпочтительно активируется после хронического самостоятельного приема кокаина, что может объяснить, почему разрушение Epac2 ослабляет самостоятельный прием кокаина, но не сахарозы».
Рисунок 5: Боковая оболочка NAc была сфокусирована. Функционально медиальная оболочка преимущественно участвует в мотивированном поведении. Некоторое обоснование сосредоточения внимания на боковой оболочке поможет.
Наш выбор латеральной оболочки NAc был обусловлен ее более плотной иннервацией от дофаминовых нейронов VTA по сравнению с другими субрегионами NAc, как показано в атласе связей мозга мыши из Института Аллена (https://connectivity.brain-map.org/) ( PMID: 246
) и наши собственные предварительные исследования.
Рисунки 7 и 8: Возможно, следует рассмотреть контрольные образцы дикого типа. В частности, помимо мышей Epac2-cKO, будет ли хемогенетическое подавление/усиление высвобождения дофамина также влиять на мышей без генетических модификаций?
Мы добавили следующее обсуждение, чтобы ответить на этот вопрос (страницы 18-19, строки 416-427) и связанные с ним вопросы, поднятые рецензентом 3.
«Мы признаем ограничения наших экспериментов DREADD. Во-первых, стимуляция Gs-DREADD использовалась для восстановления высвобождения дофамина у мышей Epac2-cKO, но не восстанавливала функцию Epac2. Дальнейшие исследования могли бы определить, может ли вирусная экспрессия Epac2 в дофаминовых нейронах VTA у мышей Epac2-cKO восстановить высвобождение дофамина и самостоятельный прием кокаина до уровня мышей WT. Во-вторых, наши исследования DREADD были ограничены экспериментами, которые были необходимы для проверки центральной гипотезы о том, что Epac2-cKO-индуцированное снижение высвобождения дофамина способствует дефициту самостоятельного приема кокаина. И мимикрия, и окклюзия могут дать представление о том, происходят ли два биологических процесса по одному и тому же механизму. Мы показали, что ингибирование hM4D(Gi) дофаминовых нейронов VTA у мышей WT имитировало эффекты Epac2-cKO при самостоятельном приеме кокаина. Дополнительным экспериментом будет определение того, снижает ли ингибирование hM4D(Gi) дофаминовых нейронов дальнейшее снижение самоконтроля кокаина у мышей Epac2-cKO или эффект ингибирования hM4D(Gi) блокируется Epac2-cKO».
Рецензент №3 (Рекомендации авторам):
1. Чтобы ответить на вопрос № 1 в Public Review, потенциальный эксперимент, который авторы могут рассмотреть, заключается в том, чтобы вернуть белок Epac2 дикого типа в мутантные нейроны VTA DA (например, посредством сверхэкспрессии вирусного гена) с последующим измерением высвобождения DA и кокаина. СА. Авторы также могут рассмотреть возможность тестирования, способствует ли хемогенная активация дофаминовых нейронов сахарозной СА, что может дать представление о природе и значении спасения. Если некоторые или все эти эксперименты неосуществимы или нецелесообразны, необходимо провести подробное обсуждение и/или обоснование для разъяснения.
Наша лаборатория не имеет прямого опыта разработки или производства вирусных векторов. Потребовалось бы значительное время, чтобы сотрудничать с Vector Core, чтобы спроектировать, упаковать и проверить сверхэкспрессию Epac2 в зависимости от типа клеток и проверить ее влияние на самостоятельное введение кокаина мышам Epac2-cKO. мРНК мышиного Epac2 (Rapgef4) содержит 3040 п.н. Вместе с регуляторными элементами и EGFP он может быть не в состоянии системы доставки, опосредованной AAV (максимум ~ 4,8 т.п.н.), но может быть доставлен с помощью лентивирусной системы. Мы очень заинтересованы в изучении этого исследования в будущем. Как упоминалось выше (вопрос 1 Public Review), проверка того, является ли хемогенная активация дофаминовых нейронов продвигает сахарозу SA может оказаться невыполнимым.
2. Чтобы рассмотреть пункт № 2 в открытом обзоре, авторы могут рассмотреть возможность повторения экспериментов по хемогенетическому ингибированию на мышах Epac2 cKO и посмотреть, блокирует ли делеция Epack2 или притупляет эффекты ингибирования высвобождения DA на кокаин SA у мутантных мышей, или предоставить более подробные сведения. обсуждения или уточнения.
Мы добавили обсуждение в исправленную рукопись (страницы 18-19, см. также наш ответ на последний вопрос рецензента 2).
3. Для решения пункта № 3 в общественном обзоре авторы должны представить временную динамику переходных изменений DA после введения DCZ и показать более репрезентативные записи сигнала DA.
Мы представили временную динамику переходных изменений DA после инъекции DCZ и показали более репрезентативные переходные процессы дофамина для записей фотометрии волокон (рис. 6).
4. Рабочая модель рисунка 9 может быть пересмотрена путем добавления некоторых промежуточных шагов между входящими пиками потенциала действия и стыковкой КА.
Мы пересмотрели модель, добавив возбуждение потенциала действия (теперь рис. 8). Тем не менее, мы удалили стыковку SV, как было рекомендовано выше (вопрос 4 Public Review).
https://doi.org/10.7554/eLife.80747.sa2
Статья и информация об авторе
Сведения об авторе
Сяоцзе Лю
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Концептуализация, Курирование данных, Формальный анализ, Валидация, Исследование, Визуализация, Методология, Написание — первоначальный проект, Администрирование проекта, Написание — обзор и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Кейси Р.
ВикстромКафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Концептуализация, Курирование данных, Получение финансирования, Исследование, Методология, Написание — первоначальный проект, Написание — обзор и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Хао Юй
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Концептуализация, Курирование данных, Формальный анализ, Исследование, Методология, Написание — первоначальный проект, Написание — обзор и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Шуай Лю
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Курирование данных, Формальный анализ, Исследование, Написание — первоначальный проект, Написание — просмотр и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Шана Терай Снарренберг
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Взнос
Курирование данных, Программное обеспечение, Исследование, Методология, Написание — обзор и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Владислав Фридман
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Курирование данных, Формальный анализ, Исследование, Написание — первоначальный проект, Написание — просмотр и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
«Этот ORCID iD идентифицирует автора этой статьи:»
0000-0002-5567-2520Ляньвэй Му
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Курирование данных, Формальный анализ, Исследование, Методология, Письмо — обзор и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Бисюань Чен
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Курирование данных, исследование, написание — обзор и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Томас Дж.
КеллиКафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Курирование данных, исследование, методология, написание — просмотр и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Дэвид Бейкер
Департамент биомедицинских наук, Университет Маркетт, Милуоки, США
Вклад
Концептуализация, Надзор, Привлечение финансирования, Написание — обзор и редактирование
Конкурирующие интересы
Конкурирующие интересы не заявлены
Цин-сон Лю
Кафедра фармакологии и токсикологии, Медицинский колледж Висконсина, Милуоки, США
Вклад
Концептуализация, Ресурсы, Надзор, Привлечение финансирования, Исследование, Написание — первоначальный проект, Администрирование проекта, Написание — обзор и редактирование
Для корреспонденции
qsliu@mcw.
eduКонкурирующие интересы
Конкурирующие интересы не заявлены
«Этот ORCID iD идентифицирует автора этой статьи:»
0000-0003-1858-1504
Финансирование
Национальный институт по борьбе со злоупотреблением наркотиками (R01DA035217)
- Цин-сон Лю
Национальный институт по борьбе со злоупотреблением наркотиками (R01DA047269)
- Цин-сон Лю
Национальный институт психического здоровья (F30Mh215536)
- Кейси Р. Викстром
Национальный институт по борьбе со злоупотреблением наркотиками (R01DA050180)
- Дэвид Бейкер
Национальный институт по борьбе со злоупотреблением наркотиками (F31DA054759)
- Владислав Фридман
Национальный институт психического здоровья (Mh221454)
- Цин-сон Лю
Спонсоры не участвовали в разработке исследования, сборе и интерпретации данных или принятии решения о представлении работы для публикации.
Благодарности
Эта работа была поддержана грантами Национального института здравоохранения F30Mh215536 (для CRV), R01DA035217 (для QSL), R01DA047269 (для QSL), R01DA050180 (для DAB и QSL) и F31DA054759.(к В.Ф.). CRV и TJK являются участниками Программы обучения ученых-медиков в MCW, которая частично поддерживается грантом на обучение от NIGMS T32-GM080202. Мы благодарим доктора Юлонга Ли (Пекинский университет, Пекин, Китай) за подарок AAV9-hSyn-GRAB DA2m и Программу снабжения лекарствами NIDA за предоставление кокаина, используемого в этих исследованиях.
Этика
Все содержание и использование животных осуществлялись в соответствии с протоколами (AUA № 2420), одобренными Институциональным комитетом по уходу и использованию животных Медицинского колледжа штата Висконсин. Все операции проводились под анестезией кетамином и ксилазином, и были приложены все усилия, чтобы свести к минимуму страдания.
Старший редактор
- Кейт М. Вассум, Калифорнийский университет, Лос-Анджелес, США
Редактор-рецензент
- Джереми Дж. Дэй, Алабамский университет в Бирмингеме, США
Рецензент
- Эрин Калипари, Университет Вандербильта, США
История публикаций
- Получено: 2 июня 2022 г.
- Препринт опубликован: 16 июня 2022 г. (просмотреть препринт)
- Принято: 21 августа 2022 г.
- Принятая рукопись опубликована: 22 августа 2022 г. (версия 1)
- Версия записи опубликована: 1 сентября 2022 г. (версия 2)
Авторское право
© 2022, Лю и др.
Эта статья распространяется в соответствии с лицензией Creative Commons Attribution License, которая разрешает неограниченное использование и распространение при условии указания оригинального автора и источника.
Показатели
Число цитирований статей, полученное путем опроса самых высоких значений из следующих источников: Crossref, PubMed Central, Scopus.
Ссылки для скачивания
Список ссылок, состоящий из двух частей, для загрузки статьи или частей статьи в различных форматах.
Открытые цитаты (ссылки для открытия цитат из этой статьи в различных онлайн-сервисах управления ссылками)
- Менделей
- ЧитатьКуб»>
Процитируйте эту статью (ссылки для загрузки цитат из этой статьи в форматах, совместимых с различными инструментами управления ссылками)
- Сяоцзе Лю
- Кейси Р Викстром
- Хао Юй
- Шуай Лю
- Шана Терай Снарренберг
- Владислав Фридман
- Ляньвэй Му
- Бисюань Чен
- Томас Дж. Келли
- Дэвид Бейкер
- Цин-сонг Лю
(2022)
Epac2 в дофаминовых нейронах среднего мозга способствует усилению приема кокаина за счет усиления высвобождения дофамина
eLife 11 :e80747.
https://doi.org/10.7554/eLife.80747
- Скачать БибТекс
- Скачать .RIS
Категории и теги
- Исследовательская статья
- Неврология
- Эпак2
- дофамин
- самоуправление
- кокаин
Исследовательский организм
- Мышь
Методы стабилизации береговой линии — Департамент охраны окружающей среды штата Нью-Йорк
Версия для печати этой веб-страницы (PDF) (1,6 МБ)
Береговая линия является ценным и важным районом. Он обеспечивает богатую и активную среду обитания для рыб и диких животных, а также очищает ливневые стоки до того, как они попадут в воду. Береговая линия обеспечивает структурную целостность кромки воды, защищая ее от эрозии. Береговая линия также дает нам расслабляющее место для рыбалки, катания на лодке и созерцания природы.
Береговая эрозия – это естественный процесс, происходящий на озерах, ручьях, реках и вдоль побережья. Это постепенное, хотя иногда и быстрое удаление наносов с береговой линии. Это вызвано рядом факторов, включая штормы, воздействие волн, дождь, лед, ветер, поверхностный сток и потерю деревьев и другой растительности. Хотя эрозия сама по себе не является вредной, когда она усиливается до такой степени, что влияет на природные ресурсы, качество воды, экосистемы и приводит к утрате собственности, это, как правило, нежелательно.
Некоторые широко практикуемые модификации береговой линии и методы борьбы с эрозией могут фактически увеличить скорость эрозии, что приводит к дорогостоящим структурным повреждениям, а также к потере имущества. Другие могут способствовать разрушению природной среды береговой линии. В этом руководстве рассматриваются проблемы, связанные с некоторыми из этих распространенных методов, и предлагаются альтернативные методы. Департамент поощряет использование «мягких» или естественных методов защиты береговой линии, а не «жестких» или структурных методов. Эти методы намного проще для окружающей среды; имитируют естественные системы, могут естественным образом взаимодействовать с экосистемой, а также экономить значительную сумму денег. Мы включили советы и информацию о различных методах стабилизации береговой линии. Имейте в виду, что для установки большинства этих методов необходимы разрешения.
Некоторые основные принципы защиты береговой линии
Чтобы наилучшим образом сохранить окружающую среду береговой линии, методы стабилизации должны соответствовать следующим основным принципам:
- Имитировать природу
Местная растительность, обычно встречающаяся на береговой линии, укрепляет ее структурную целостность и предотвращает разрушение земли. Глубокие корни этих растений связывают землю воедино, а их листва и ветви защищают от эрозии, вызванной дождями и ветрами. Удаление этих растений может привести к тому, что берег ослабнет и легко рухнет в воду. - Держите склоны пологими
Пологий уклон естественной береговой линии поглощает энергию волн. Крутой эрозионный склон или подпорная стенка позволяет волнам разбиваться о берег, резко увеличивая эрозию и вызывая эту энергию волн, наносящую ущерб прилегающим береговым линиям. - По возможности используйте «мягкую броню».
Под «мягкой защитой» мы подразумеваем живые растения, бревна, корневые пыжи, растительные маты и другие методы, которые устраняют или уменьшают потребность в «жесткой защите», такой как каменная наброска, каменные блоки, шпунт или другие твердые материалы. материалы. Мягкая броня живая и поэтому может адаптироваться к изменениям окружающей среды, а также размножаться и размножаться. Он также обеспечивает среду обитания для рыб и диких животных. Растительность можно подстригать, чтобы не загораживать обзор — ведь именно поэтому многие из нас предпочитают жить у воды! - Смешайте это
Независимо от типа естественной береговой линии, вы, несомненно, увидите большое разнообразие материалов: живые деревья, сухие ветки, пни, камни разных форм и размеров, ил, песок, рогоз, травы, цветущие растения и т. д. разнообразие, вы можете сохранить или воспроизвести естественную ценность береговой линии и получить эффективную, устойчивую и приятную для глаз береговую линию. Работа с этими природными и местными материалами также может значительно снизить стоимость проекта. В конце концов, сочетание методов может привести к лучшему проекту, уникально подходящему для вашей ситуации. Если ваш район уже застроен, рекомендуется поискать примеры более естественного состояния на близлежащих незастроенных участках береговой линии. - Небольшой и простой.
В некоторых случаях эрозии может подвергаться только часть вашей береговой линии. В этом случае может быть достаточно небольшого или смешанного проекта с каменной наброской и растительностью. Имейте в виду, что здоровые деревья часто являются краеугольным камнем стабильной береговой линии.
Как естественная береговая линия может защитить от эрозии?
В естественном состоянии береговая линия идеально защищена от эрозии.
Возьмем пример ниже:
Местная растительность, обычно встречающаяся в прибрежной зоне, укрепляет ее структурную целостность и предотвращает разрушение земли. Глубокие корни этих растений связывают землю воедино, а их листва и ветви уменьшают эрозию, вызванную дождями и ветрами. Очистка этих растений приводит к тому, что берег ослабевает и легко рушится в воду.
Пологий уклон естественной береговой линии образовался для поглощения энергии волн. Крутой эрозионный склон или подпорная стенка позволяет волнам разбиваться о берег, резко увеличивая эрозию.
Это та же береговая линия после значительной эрозии:
Хотя это может не совсем точно отражать вашу береговую линию, это изображение представляет общие последствия эрозии.
Как прибрежная зона обеспечивает среду обитания для рыб и диких животных?
Естественная береговая линия:
- Рыбы и лягушки часто нерестятся в иле на дне берега и в прибрежной зоне. Прибрежная зона находится сразу за береговой линией по направлению к воде.
- Растительность обеспечивает места гнездования птиц и пищу для насекомых, водоплавающих птиц и водных млекопитающих.
- Упавшие бревна и ветки служат убежищем и охотничьими угодьями для рыб и млекопитающих, а черепахи используют их для принятия солнечных ванн.
- Естественная растительность береговой линии действует как фильтр, предотвращая попадание отложений и ненужных питательных веществ в водоем. Этот сток приводит к ухудшению качества воды и нарушает баланс, необходимый для здоровой среды обитания на береговой линии. В случае газонов этот сток может включать удобрения, пестициды, обрезки газонов и отходы домашних животных. Гусей привлекают газоны, и их отходы могут увеличить этот сток.
- Черепахи и земноводные имеют свободный доступ к суше, а подпорные стены (см. ниже) блокируют этот доступ, предотвращая спаривание и нерест.
Это та же самая береговая линия после того, как землевладелец установил подпорную стенку:
Эффекты хорошо видны, так как пышная экосистема быстро разрушается.
Вредные изменения береговой линии
Следующие виды изменений и защиты береговой линии приводят к усилению эрозии, структурным повреждениям и разрушению экосистемы береговой линии:
Переборки и подпорные стенки
Обычно естественный пологий уклон поглощает энергию волны. Переборки и подпорные стены заставляют волны разбиваться о берег. Большая часть энергии этих волн направляется вниз в воду у основания стены. Грязь или «подложка» медленно выкапывается из-под основания стены, в конечном итоге заставляя ее наклоняться, а затем падать в воду. Кроме того, вода, стекающая с возвышенности, скапливается за стеной, толкая стену сзади, особенно во время циклов замерзания-оттаивания. Строительство переборки или подпорной стены также может увеличить скорость эрозии соседних участков.
Влияние переборки на соседние
непереборочное имущество
Предоставлено Службой охраны рыбных ресурсов и дикой природы США
С точки зрения охраны окружающей среды подпорные стенки являются наиболее разрушительным методом стабилизации. С помощью подпорных стен территория с растительностью, которая обычно обеспечивает укрытие, места для кормления, места размножения и гнездования, расчищается и застраивается, быстро и полностью разрушая экосистему.
Разрушение подпорной стены
Мало того, что подпорные стены являются самым дорогим и вредным для окружающей среды вариантом, но когда они в конце концов рухнут, вы останетесь с ошеломляющим счетом за ремонт. Это особенно проблематично в тех случаях, когда такие конструкции, как дома, гаражи и т. д., строятся вплотную к стене. Если стена рухнет, вы можете обнаружить, что сопутствующая конструкция рушится вместе с ней. Каким бы обширным он ни был, обрушение подпорной стены неизбежно.
Очень обширная, но неудавшаяся подпорная стена
Если у вас уже установлена подпорная стена, вы можете полностью удалить стену и переделать участок одним из рекомендуемых нами методов. Однако в некоторых местах вы не сможете использовать один из этих вариантов. В этом случае Департамент может порекомендовать вам построить стену дальше (к суше) от существующей стены, а не строить новую стену дальше в воду.
Постоянные доки
Доки могут прерывать поток наносов вдоль берега и вызывать неестественные отложения, влияющие на скорость эрозии. Они также могут блокировать солнечный свет, необходимый для здоровой растительности. По этим и другим причинам лучше минимизировать размер и количество доков. Временные и сезонные доки являются оптимальным компромиссом, требующим небольшого количества труда для сезонной установки.
Искусственные песчаные пляжи
Важно сохранить естественный состав субстрата. Многие люди либо сбрасывают песок, чтобы создать пляжную зону, либо убирают естественную растительность. Песчаные пляжи подвержены эрозии и убивают растительность под ними. Песок легко смывается отступающими волнами. Если желательна песчаная поверхность, ее следует расположить вдали от зоны воздействия волн, с широкой буферной полосой с растительностью. Удаление местной растительности и слишком большого количества упавших бревен и веток увеличивает скорость эрозии и наносит вред экосистеме водоема. Корни растительности скрепляют берег и служат пищей и убежищем для водных млекопитающих, птиц, черепах и насекомых.
Газоны и химикаты для газонов
Ухоженный газон, размытый волнами
Ухоженные газоны лишены растительности, которая обычно предотвращает эрозию береговой линии. Трава, используемая для этих газонов, не имеет глубокой корневой системы, которая необходима для стабильной береговой линии. Сочетание волн и погоды начнет разъедать газон, заставляя его рассыпаться и рухнуть в воду.
Химикаты для газонов, такие как удобрения и гербициды, могут попадать в водоем и влиять на его химический баланс, способствуя неприглядному и нездоровому цветению водорослей или, в тяжелых случаях, гибели рыбы.
Чтобы предотвратить эти проблемы: Либо замените газон местной растительностью, либо создайте «буферную зону», чтобы отделить газон от воды.
Как создать буферную зону:
Буферная зона представляет собой полосу растительности у кромки воды. Чем шире буферная зона, тем лучше, но буфера, простирающегося не менее чем на 50 футов назад, должно быть достаточно, хотя для озер с холодной водой потребуется не менее 100 футов. Конечно, даже самый маленький буфер лучше, чем ничего. Самый простой способ создать буферную зону — перестать косить полосу газона у кромки воды. Затем в этой области должна начать возвращаться местная растительность. Вы можете посадить определенные виды растений либо из-за их внешнего вида, либо из-за их эффективности в стабилизации берега. Дополните траву глубоко укоренившейся древесной растительностью. Поскольку местные виды различаются в разных районах штата, мы рекомендуем вам проконсультироваться с местной службой охраны почв и водных ресурсов для получения предложений.
Пешеходные дорожки или дороги
Если вдоль берега проложены пешеходные дорожки или дороги, ливневые стоки резко увеличатся, что значительно усугубит эрозию. Обычно земля впитывает дождевую воду. Как и в случае с газонами, любые пешеходные дорожки или дороги должны быть должным образом защищены от воды, как описано выше.
Рекомендуемые методы защиты береговой линии
Предоставлено Клэр Прайн, DEC Region 8
Мы рекомендуем использовать «более мягкие» подходы для защиты береговой линии. Эти методы могут быть более экономичными (с более низкими затратами на техническое обслуживание), более долговечными и эластичными, эстетически привлекательными и экологически безопасными по сравнению с обычно используемыми «жесткими» или структурными методами. Некоторые землевладельцы также предпочитают эти методы, потому что они помогают береговой линии гармонировать с окружающей природой.
Обычно подрядчики рекомендуют установить бетонную подпорную стену. Хотя это может показаться практичным решением, мы настоятельно рекомендуем вам рассмотреть один из рекомендуемых нами методов. Подпорные стенки являются не только самым дорогим вариантом, но и имеют склонность к обрушению, что требует капитального ремонта. Они также оказывают негативное влияние на окружающую среду.
Восстановление растительности
Где применяется: Работы по восстановлению растительности на газонах или голых береговых линиях с эрозией от слабой до умеренной. Это не для береговых линий с обширными повреждениями или сильными волнами.
Основная идея: Этот метод включает повторную посадку местной растительности, которая естественным образом стабилизирует береговую линию. Глубокие корни этих растений плотно связывают землю внизу, эффективно защищая береговую линию от эрозии.
Стоимость: Низкая
Сложность: Легкая, может быть выполнена землевладельцем
С голыми берегами:
В случаях, когда береговая линия оголена, растительность придется высаживать самостоятельно. Сажайте поздней осенью или ранней весной для большей успешности. Это поглотит первоначальное волновое воздействие.
Live Staking
Где работает: Склоны с легкой эрозией; может использоваться в сочетании с другими методами для участков с более сильной эрозией.
Основная идея: Возьмите черенки древесных растений (живые колья), такие как ива и кизил, и вбейте их в грязь или субстрат эродированного участка. Они пускают корни и растут. Обычно это лучше делать ранней весной или поздней зимой.
Стоимость: Низкая
Сложность: Легко, может выполнить землевладелец
Предоставлено Министерством сельского хозяйства США — Роббин Б. Сотир и
Партнеры
1. Сбор и подготовка ставок
Колья должны быть срезаны (длиной 1 фут или более) ножницами из спящих, зрелых стеблей и должны быть использованы в течение 8-10 дней. Обрежьте боковые ветки, не повредив кору, и срежьте нижнюю часть кола под углом.
2. Забивные стойки
Используя безударный молоток, осторожно вбейте ивовые или кизиловые колья в землю под прямым углом к склону (см. схему). Если почва слишком уплотнена, предварительно сделайте отверстие стальным стержнем. Колья должны быть установлены плотно, примерно на 70% закопаны, а на 30% открыты. Разместите колья в виде треугольника на расстоянии от 2 до 3 футов друг от друга, максимум 4 кола на ярд. Геотекстильная ткань или джутовая сетка необязательны, но могут понадобиться для предотвращения дальнейшей эрозии, пока живые колышки не начнут расти. Просто разрежьте и положите ткань на участок, прежде чем вбивать какие-либо колья. Закрепите ткань, погрузив ее конец в субстрат за кольями. Rip Rap на носке необязателен.
Contour Wattling (Live Fascines)
Предоставлено USDA — NRCS EFH Chapter-16
Где это работает: Склоны с легкой эрозией
Основная идея: Положите живые пучки стеблей и веток в траншеи на береговой линии и засыпьте их почвой. Они удерживаются на месте как деревянными, так и живыми кольями. Они пускают корни и растут.
Стоимость: Низкая
Сложность: Легко умерить в зависимости от необходимого уровня подготовки площадки.
Предоставлено USDA — NRCS EFH Chapter-16
1. Соберите и подготовьте пакеты
Связки сделаны из свежих черенков растений. Они должны быть прямыми, диаметром от 6 до 8 дюймов и длиной от 5 до 30 футов в зависимости от условий на участке. Черенки растений должны различаться по возрасту, размеру и виду, при этом растущие кончики должны быть обращены в одном направлении. Особенно хорошо для этого применения подходят ивы и кизилы. Боковые ветки черенков обрезать не нужно. Связанные вместе пучки должны быть от 6 до 8 дюймов в диаметре. Вы можете использовать конопляный или манильский связующий шпагат или другую разлагающуюся ткань, чтобы связать пучки, располагая завязки на расстоянии около фута друг от друга.
Предоставлено Министерством сельского хозяйства США — Роббин Б. Сотир и
Ассоциированные сотрудники
2. Рытье траншей и укладка связок
Выкапывайте траншеи, начиная с основания склона и продолжая вверх по береговой линии, располагая их на расстоянии около 3 футов друг от друга. Траншеи должны соответствовать ширине ваших связок, а их глубина должна составлять примерно половину диаметра пачки, т. е. 3-4 дюйма. Положите свои узлы в траншеи.
Предоставлено Министерством сельского хозяйства США — Robbin B. Sotir & Associates
3. Безопасные пакеты
Закрепите связки на месте, вбив деревянные колья длиной от 2 до 3 футов через связку и в почву под ней. Разместите колышки на расстоянии 2–3 фута друг от друга и убедитесь, что из верхней части связки торчит не менее 2–3 дюймов колышка. Затем засыпьте пучки почвой, но оставьте небольшую часть верхушки пучка открытой. Несколько больших живых кольев из черной ивы можно разместить вместе с мертвыми кольями, чтобы закрепить связку, помогая восстановить растительность на склоне.
Предоставлено Министерством сельского хозяйства США — NRCS EFH Chapter-16
4. (Необязательно) Укладка соломы, мульчи или ткани
На более пологих склонах между связками укладывайте солому или мульчу. На более крутых склонах постелите джутовую или кокосовую (кокосовое волокно) ткань. Ткань должна проходить под пучками, а пучки должны быть закреплены в ткани. Ткань должна быть установлена до шага 3.
Наслоение кистью
Где работает: На сильно эродированных склонах
Основная идея: В склоне выкапываются лунки, в которые вставляются черенки растений под углом и растут наружу, а корни врастают в склон.
Стоимость: Низкая
Сложность: Средняя
1. Обрезка веток
Срежьте свежие спящие ветки (длиной до 12 футов и диаметром от 1/4 до 1 дюйма) выбранных вами видов, предпочтительно ивы или кизила. Обрежьте боковые ветки.
2. Выкопайте первую скамейку
Выкопайте первую скамейку в нижней части склона. Скамья должна быть вкопана в склон под углом от 10 до 25 градусов от горизонтали (см. схему)
3. Поместите черенки
На каждой скамье должно быть несколько слоев черенков, каждый слой должен быть толщиной около дюйма. Положите первый слой так, чтобы нижние концы касались задней части выкопанного участка, а кончики были направлены вверх от склона. Черенки должны быть перекрещены. Покройте этот слой грязью и приступайте к следующему. Повторяйте этот процесс, пока не будет достигнуто желаемое количество слоев. Затем выкопайте следующую скамейку и повторите шаги 1-3.
4. Укладка мульчи или соломы
Положите мульчу или солому на открытую почву между скамейками.
Матрац из щеток
Где применяется: На сильно эродированных склонах
Основная идея: Это включает создание «матраса из щеток» из живых обрезков растений, который лежит на эродированном участке. Он укоренится и разрастется.
Стоимость: Низкая
Сложность: Средняя
1. Выберите растение
Матрасы из щеток изготавливаются из любого древесного растения, у которого пускают корни из стебля. В основном люди используют иву, но также подойдут некоторые виды кизила и калины. Растения должны быть в возрасте от 2 до 3 лет, гибкими и длиной от 5 до 10 футов. Диаметр ветвей должен быть от ½ до 1½ дюймов.
2. Подготовка откоса
Склон, на который вы будете укладывать матрас, должен быть ровным, чтобы каждая часть матраса соприкасалась с почвой. Площадь должна быть от 5 до 18 футов в длину. Убедитесь, что почва достаточно рыхлая, чтобы матрац мог укорениться.
3. Выкопать траншею
Выкопайте траншею по длине носка склона, где будет заканчиваться матрац из кистей. Здесь будет заправлен конец матраса. НЕОБЯЗАТЕЛЬНО, но рекомендуется: Создайте живой фашин (см. раздел «Контурное плетение» выше) и положите его в траншею.
4. Ставить ставки
Рекомендуется использовать комбинацию живых и мертвых ставок. Ставки должны быть толщиной от 2 до 4 дюймов и длиной от 24 до 36 дюймов. Вбейте их в склон рядами на расстоянии 18 дюймов друг от друга снизу вверх по склону (см. схему). Если вы решили включить живой фашин, колья в нижней части рядов должны быть вставлены прямо через фашин.
5. Сложить ветки
Теперь положите ветки между рядами кольев, создавая слой толщиной от 2 до 4 дюймов.
6. Привяжите их
Выбирайте проволоку или шпагат. Если используете проволоку, используйте калибр 16. Для шпагата используйте кокосовую стружку толщиной от 1/5 до ½ дюйма с прочностью на разрыв от 70 до 100 фунтов. Проволока или бечевка проходит по диагонали через колышки (см. схему) и привязывается к каждому кольцу с помощью зубчатой петли. Rip Rap на носке необязателен.
Предоставлено Министерством сельского хозяйства США
.
Предоставлено Министерством сельского хозяйства США
.
Предоставлено Министерством сельского хозяйства США — NRCS EFH Chapter-16
Противоэрозионное покрытие
Основная идея: Лист специальной трехмерной биоразлагаемой геотекстильной ткани для предотвращения эрозии укладывается на открытый склон береговой линии. Семена травы сажают по всему покрытию, а затем засыпают землей. Трава переплетается с матом или покрывалом и стабилизирует берег.
Где работает: Умеренные уклоны до 1 вертикали до 2 горизонталей вдоль проезжей части или на склонах вдоль водотоков.
Стоимость: От низкой до умеренной в зависимости от площади покрытия и необходимой подготовки площадки.
Сложность: От умеренной до сложной, в зависимости от необходимой подготовки склона и характеристик площадки.
1. Подготовка откоса
Удалите все комки почвы и камни. Подготовленная площадь должна точно соответствовать размерам проекта. Необязательно — разбросайте семена под областью, на которую вы будете укладывать коврик.
2. Противоэрозионные покрытия поставляются в рулонах. Постелите одеяла
Каждое одеяло обычно имеет ширину от 3 до 4 футов. Начиная сверху, скатывайте каждое одеяло вниз по склону. Каждое одеяло должно перекрывать следующее примерно на 3-4 дюйма.
3. Скрепите одеяло скобами
Вбейте специальные скобы для почвы по центру каждого одеяла на расстоянии от 3 до 5 футов друг от друга.
4. Рассыпьте почву поверх одеяла
Тщательно распределите слой почвы толщиной от 1/2 до 3/4 дюймов на коврике, это поможет семенам укорениться.
5. Разбросайте семена
Разложите семена по коврику. Распространение семян под матом — еще один рекомендуемый вариант. Сделайте это перед укладкой коврика.
Методы защиты береговой линии, рекомендуемые при определенных условиях
Следующие методы включают жесткую броню и должны использоваться только в том случае, если описанные выше методы не будут работать на вашей собственности. Департамент предпочитает более мягкие методы, описанные выше, поскольку они лучше предотвращают эрозию и поддерживают здоровую окружающую среду на береговой линии. Если действительно требуется жесткое армирование, наклонная каменная наброска или комбинация растительности с каменной наброской обеспечивают лучшую защиту береговой линии, чем вертикальные стены из цемента или блоков, и при этом обеспечивают некоторую среду обитания для диких животных. Все эти методы потребуют разрешения Департамента.
Предоставлено Министерством сельского хозяйства США — NRCS EFH Chapter-16
Stone Rip-Rap
Где работает: Береговая линия, где подстилающий грунт стабилен.
Основная идея: Слой камней укладывается вдоль склона или берега и предотвращает эрозию, вызванную воздействием волн.
Стоимость: От средней до высокой
Сложность: Средняя
1. Подготовьте склон
Методы вегетативной стабилизации предпочтительнее на более плоских склонах, и использование каменной каменной наброски, как правило, должно быть ограничено склонами более чем от одной вертикальной до трех горизонтальных, где методы вегетативной стабилизации не столь эффективны. Склон должен быть градуирован максимум не более чем на один фут по вертикали на каждые 1,5 фута горизонтального расстояния. (В некоторых регионах DEC требуется уклон 1:2 в зависимости от условий участка.) Почва должна быть стабильной. Если вам нужно заполнить площадь, используйте камни размером менее 6 дюймов. Перед укладкой каменной наброски убедитесь, что склон плотно утрамбован.
2. Укладка каменной крошки
Каменная наброска – это твердый добытый в карьере камень, такой как куски известняка или гранита, обычно длиной 12–18 дюймов, имеющие не менее двух изломов. Эти камни сцепляются друг с другом. Не используйте закругленные камни, потому что они будут скользить вниз на склоне. Должно быть два слоя. Первый слой называется «фильтрующий слой» Камни должны быть не больше 3 дюймов в диаметре. Если вы решите использовать фильтровальную ткань, поместите ее под фильтрующий слой. Верхний слой или «броневой слой» и принимает на себя первоначальное воздействие волн. Если площадь достаточно велика, вам, возможно, придется использовать кран или самосвал для сброса камней. Если проект достаточно мал, вы можете это сделать. вручную
Примечание: Если на эродированном склоне есть живые деревья или другая значительная местная растительность, мы настоятельно рекомендуем оставить их на месте, а вокруг них уложить каменную наброску, стараясь не повредить кору или растительность.
Предоставлено Министерством сельского хозяйства США — NRCS EFH Chapter-16
Rip-Rap с растительностью (система совместной посадки)
Где применяется: Водные пути или внутренние озера с устойчивым подстилающим грунтом.
Основная идея: Это сочетание живого сталкинга и рип-рэпа. Каменная насыпь предотвращает размывание берега волнами, в то время как корни растений связывают землю внизу. Растения покрывают скалы, создавая тень для рыб и диких животных и, в конечном итоге, создавая очень хорошее место для рыбной ловли. Некоторые землевладельцы также предпочитают более естественный вид береговой линии.
Стоимость: От средней до высокой. Гораздо дешевле, если рип-рэп уже на месте.
Сложность: Высокая
1. Подготовить склон
Наклон должен быть 2 к 1 (от горизонтального к вертикальному). Накройте это место фильтрующей тканью или джутовой сеткой. Распределите камни по ткани, стараясь не повредить ее.
2. Подготовьте живые ставки
Колья должны быть обрезаны достаточно долго, чтобы их можно было вонзить в грязь под камнями, при этом растущие кончики должны выступать над камнями. Срезают ножницами из спящих, зрелых стеблей и должны быть использованы в течение 8-10 дней. Обрежьте боковые ветки, не повредив кору, и срежьте нижнюю часть кола под углом.
3. Вставьте живые ставки
Их следует вставлять перпендикулярно откосу. Растущие кончики должны быть обращены вверх, выступая сквозь камни и над ними. Вы можете использовать стальной стержень (арматурный стержень), чтобы проткнуть ткань и создать отверстие в почве под ней. Используйте ударный молоток, чтобы вбить колья в почву. На квадратный метр должно приходиться от двух до четырех кольев.
NRCS EFH Глава-16
Предоставлено USDA
Матрас из габиона с растительностью
Основная идея: Габионный матрац представляет собой удлиненную клетку в форме матраца, заполненную камнями. Габионные матрацы с растительностью включали ветки или черенки, вставленные сквозь камни в клетку.
Где это работает: Умеренные уклоны для защиты от воздействия волн, льда и поверхностной эрозии.
Стоимость: Высокая
Сложность: Высокая. Мы рекомендуем вам не пытаться сделать это самостоятельно. Требуется профессиональный подрядчик, тяжелая техника и инженер.
Озелененные грядки (Живые грядки)
Основная идея: Переплетенные деревянные доски действуют как своего рода живая подпорная стена, но с меньшим воздействием на окружающую среду. Между досками высаживают растительность. Это не очень хорошо работает на высоких берегах с сильными волнами.
Где это работает: Склоны без растительности с большим количеством обратной засыпки и небольшим воздействием волн
Стоимость: От средней до высокой
Сложность: Средняя. Мы рекомендуем вам не пытаться сделать это самостоятельно. Требуется профессиональный подрядчик, тяжелая техника и инженер.
Источники
- «Справочник по стабилизации береговой линии» Северо-западная региональная комиссия по планированию. Сент-Олбанс, Вермонт.
- Полевой инженерный справочник NRCS Министерства сельского хозяйства США, часть 650, глава 16, Защита берега ручья и береговой линии. Декабрь 1996.
Строительство лодочных рамп | Вирджиния DWR
- Критерии площадки
- Разрешения
- Дизайн
- Методы строительства
- Материалы
Строительство лодочных аппарелей, независимо от того, строите ли вы аппарели для личного пользования или, в случае Департамента охоты и рыболовства во внутренних водах (DGIF), общественного пользования, многие аспекты, которые необходимо учитывать, одинаковы. Некоторыми из этих вещей являются критерии места, разрешения, дизайн, методы строительства и материалы, а также тип и размер лодок, которые будут спущены на воду.
Первое, что следует учитывать при выборе аппарели, — это тип и размер лодок, которые необходимо спускать на воду и поднимать. Очевидно, что выбор места и проектирование рампы для спуска каноэ или лодки с крышей на мелководье будет сильно отличаться от выбора места и проектирования рампы для спуска больших лодок с прицепом в приливной воде. Если вы проектируете пандус для личного пользования, довольно легко выбрать конструкцию, подходящую для лодок и буксирующих транспортных средств, которыми вы владеете или могли бы владеть. Выбрать дизайн, который подойдет нескольким лодочникам или широкой публике, сложнее. DGIF пытается спроектировать рампы, подходящие для большинства лодок в районе предполагаемой рампы, зная, что мы не можем удовлетворить потребности каждого лодочника.
В этой статье кратко рассматриваются критерии места, разрешения, дизайн, методы строительства и материалы для типичных аппарелей для спуска и подъема лодок в диапазоне 12-20 футов, которые обычно используются для спортивной рыбалки и прогулочного катания на моторных лодках. В этой статье представлен ряд пунктов для вашего рассмотрения, если вы думаете о строительстве лодочной рампы, но она не предназначена для использования в качестве руководства по строительству лодочной рампы.
Критерии площадки
При выборе и оценке площадки в качестве потенциальной площадки для лодочной рампы необходимо учитывать ее доступность, близость к другим лодочным рампам, глубину воды, степень заиления и полезную площадь земли (парковка, радиус поворота и т. д.). .).
Доступность площадки
Одним из наиболее очевидных соображений при выборе площадки является ее близость к существующим дорогам и другим пандусам. Строительство и содержание дорог требует больших затрат, поэтому чем ближе участок находится к обслуживаемой дороге, тем лучше. Кроме того, DGIF предпочитает не конкурировать с частными платными пандусами, если они могут удовлетворить общественный спрос. Тем, кто думает о строительстве собственного пандуса, было бы неплохо рассмотреть возможность использования существующих пандусов, если они доступны, и даже разделить расходы на техническое обслуживание с владельцем.
Глубина воды
Глубина воды должна быть не менее трех футов в конце пандуса во время среднего маловодья, хотя более желательна глубина четыре фута. Если прицепы с приводом будут использоваться для спуска и подъема лодок (мощная погрузка), рассмотрите возможность расширения пандуса на глубину до пяти футов или установки каменной наброски в конце пандуса. Другой альтернативой является увеличение уклона пандуса на последних 10-15 футах, чтобы конец пандуса находился в более глубокой воде или был врыт в дно, чтобы защитить конец пандуса. Если конец аппарели не защищен, промывка опоры, создаваемая силовой нагрузкой, вызовет эрозию отверстия в конце аппарели, что вызовет резкий обрыв и может подорвать конец аппарели. Если затем колеса прицепа отъезжают от конца аппарели, которая имеет спуск, прицеп может повиснуть на конце аппарели, что приведет к повреждению прицепа, когда колеса снова втянутся на аппарель. Большинство старых аппарелей DGIF не рассчитаны на прицепы с приводом, и мы часто добавляем каменную наброску в конце аппарелей в качестве временного ремонта проблем, вызванных промывкой опор.
Скорость заиливания
Еще одним соображением о глубине воды является вероятность того, что глубина воды не будет оставаться постоянной. Речные русла смещаются из стороны в сторону и могут подрезать конец вашего пандуса, в результате чего конец сломается. То же самое относится и к каналам в приливных зонах, особенно когда участок находится в точке, которая сужает более крупный залив. Как раз обратное верно для участков позади небольших бухт, вблизи устьев ручьев или на длинных песчаных пляжах. Эти места часто являются областями активных отложений, где ил или песок могут покрыть вашу рампу. Дноуглубительные работы и ремонтные дноуглубительные работы для получения глубоководных участков являются дорогостоящими, могут неблагоприятно воздействовать на окружающую среду и часто сложны, поскольку должны быть расположены подходящие места для размещения извлеченного материала. Если можно избежать дноуглубительных работ, сделайте это!
Размер
Размер пандуса и парковочной зоны зависит от предполагаемого использования. Наше общее практическое правило заключается в том, что на одной стартовой полосе должно быть около 30-35 парковочных мест для легковых автомобилей. Большинство наших рамп имеют суточную оборачиваемость от 2,0 до 2,5, поэтому каждая пусковая полоса и 30-35 парковочных мест рассчитаны примерно на 80 пусков в сутки. Если предполагается не более 80 пусков в день, достаточно одной пусковой полосы. Более 80 пусков оправдывают дополнительные пусковые полосы и парковочные места. При принятии решения о том, сколько парковочных мест может быть предоставлено на участке земли, следует помнить, что парковочное место для автомобиля с прицепом должно быть не менее 10 футов в ширину и 40 футов в длину с достаточным пространством для маневра, чтобы выстроиться в линию, въехать и выйти из него. парковочное место.
Предупреждение относительно размеров лодочной рампы: не следует недооценивать необходимый участок земли. Помните, что большие заболоченные территории нельзя трогать. Кроме того, в начале рампы необходим радиус поворота 30 или 40 футов и площадка для подготовки, и независимо от того, насколько большими вы сделаете общественные объекты, все равно будет какой-то спокойный, солнечный, теплый выходной день, когда этого будет недостаточно. Если возможно, зарезервируйте места для расширения и переполненной парковки.
Разрешения
После того, как вы нашли место для лодочной рампы, узнайте достаточно о том, как вы хотите ее построить, чтобы подготовить эскиз. Следующим шагом является подача заявки на получение необходимых разрешений. Комиссия по морским ресурсам Вирджинии, Инженерный корпус, Государственный совет по контролю за водными ресурсами и местный совет по водно-болотным угодьям могут потребовать разрешения. В некоторых населенных пунктах также требуется разрешение на строительство. Чтобы узнать, какие разрешения потребуются, свяжитесь с Комиссией по морским ресурсам Вирджинии по адресу 2600 Washington Avenue, Newport News, Virginia 23607 (тел. 757-247-2200) и с местным строительным инспектором.
Дизайн
Дизайн участка, пандуса и пирса будет иметь большое влияние на затраты на строительство и техническое обслуживание, а также на удобство использования сооружений. Потратьте время, необходимое для получения прочной, экономичной и функциональной конструкции.
План площадки
Вероятно, самая распространенная проблема при посадке лодок заключается в том, что пандус, пирс, радиус поворота и/или парковка слишком малы. Обеспечьте достаточное пространство для размещения буксирующего транспортного средства и прицепа относительно аппарели, а также достаточно места для плацдарма, чтобы подготовить лодку к спуску на воду. Если возможно, разместите пирс с той же стороны пандуса, что и водитель. Это даст водителю лучший обзор, когда он едет с прицепом вдоль пирса. Желательно одностороннее движение на стоянке и в зоне остановок, чтобы уменьшить заторы. Угловую парковку обычно легче выполнить, чем другие планы парковки, и она уменьшает необходимую ширину полос движения на стоянке. Парковочные места и схемы движения должны быть четко обозначены, чтобы уменьшить заторы на дорогах и обеспечить максимальную эффективность имеющихся парковочных мест. Уклон в один процент (минимум) поперек парковки и промежуточной площадки помогает предотвратить скопление воды на этих участках, но вызывает медленный сток, что снижает эрозию на необработанных поверхностях. Уклон должен быть по возможности направлен в сторону от пандуса, чтобы предотвратить попадание гравия, песка и т. д. на пандус. Если должны быть построены две спусковые полосы, один пирс между полосами может обслуживать обе полосы и снижать затраты, а также гарантирует, что один лодочник не сможет связать обе полосы одновременно. Для объектов с высокой интенсивностью использования двухполосная рампа с L-образными опорами с обеих сторон поможет уменьшить заторы во время пикового запуска и подъема. В центре рампы должна быть нанесена линия, чтобы помочь лодочникам оставаться на своей стороне рампы.
Конструкция пандуса
Как правило, пандусы должны иметь уклон 12–15 %, при этом бетон должен заходить в зону разворота и останова достаточно далеко, чтобы колеса прицепа могли преодолеть наклонный участок пандуса до того, как тяговые колеса тягача оторвутся от бетона. . В противном случае тяговые колеса тягача могут провернуться и зацепиться за поверхность, когда прицеп и катер поднимаются по аппарели. Пандусы шириной 16 футов предпочтительны для широкой публики, хотя многие существующие 12-футовые пандусы оказались удовлетворительными для малоиспользуемых объектов. Пандусы, установленные в текущих реках, должны входить в реку под углом вниз по течению, чтобы уменьшить боковое давление на лодку, когда она помещается на прицеп или снимается с него. Также пандус, поставленный под углом, обычно меньше накапливает ил после периода половодья. Если необходимо сделать выемку в берегу реки, откосы должны быть как можно дальше назад, чтобы уменьшить количество стоячей воды, оставшейся в выемке во время затопления, тем самым уменьшив количество ила, отложившегося на пандусе. Обеспечьте стабилизированные канавы с каждой стороны пандуса для стока во время сильных дождей.
Конструкция пирса
Если пирс необходим для спуска на воду и подъема лодок, пристальное внимание к мелким деталям конструкции пирса может сэкономить много головной боли и затрат на техническое обслуживание. Хотя пирсы могут быть изготовлены из других материалов, кроме дерева, здесь будет обсуждаться только дерево. Помните, что конструкция, в которой используются пиломатериалы стандартной длины, будет более экономичной.
Опоры могут быть как плавучими, так и стационарными. Наш опыт работы с плавучими доками показывает, что их трудно удерживать на месте, если вдоль бортов не предусмотрены сваи, чтобы пирс мог подниматься и опускаться. Металлические бочки не должны использоваться для флотации. Пластмассовые пенопластовые поплавки являются хорошим выбором, когда необходима плавучесть.
Примечание: Звучит хорошо, но непрактично, в глинистой или илистой почве это также приведет к нестабильной свае. Повреждение льдом необходимо учитывать на всех пандусах в Вирджинии. Ледоход на реках и приливах может быть особенно разрушительным. Если ожидается подвижный лед, прочность материалов должна быть выше, а конструкция усилена.
Пирсы могут повредить лодки, а лодки могут повредить пирсы. Все головки болтов и гвозди, которые могут соприкасаться с лодкой, должны быть утоплены. Следует позаботиться о том, чтобы на пирсе не было «губ», где лодка может дрейфовать и быть захваченной, если вода поднимется. Чтобы лодки не зацепились за пирс, могут понадобиться рубры.
При проектировании пирса необходимо учитывать безопасность лодочников. Настил должен перекрывать внешние стрингеры не более чем на два дюйма, чтобы настил не перевернулся в случае, если он ослабнет и кто-то наступит на его конец. Многие лодочники совершали быстрые поездки в воду или свои лодки, наступая на незакрепленные доски, когда настил значительно перекрывал внешний стрингер. Перекрытие настила на два дюйма также предотвращает расщепление большинства концов настила, когда он прибит к стрингерам пирса. Там, где поверхность воды будет значительно колебаться, должны быть установлены трапы, чтобы облегчить лодочникам посадку в лодки в периоды низкого уровня воды.
Один из аксессуаров, который хорош на пирсе, — это бордюр. Бордюр обеспечивает хорошую опору для рук при подъеме и выходе из лодок, является отличным местом для привязки лодки к пирсу и служит отбойником, помогающим предотвратить падение оборудования с пирса. Если шипы все же желательны, их можно разместить поверх бордюра, чтобы уменьшить вероятность того, что кто-то споткнется о них.
Методы строительства
Существует несколько способов построить бетонную лодочную рампу на участке. Также доступны сборные бетонные плиты, подходящие для использования в качестве пандуса. Разнообразие сужается в методах строительства опор и в основном ограничивается тем, как устанавливаются сваи.
Строительство пандуса
Хотя бетон можно смешивать для укладки (заливки) через воду, контроль качества обычно страдает, а окончательные результаты неудовлетворительны. Этот метод обычно не разрешен разрешительными органами и не будет обсуждаться в этой статье. Наилучший способ сооружения подводной части аппарели – перекрытие рампы, откачка воды, установка (заливка) и отделка аппарели всухую (монолитная). Этот метод обеспечивает наилучший горизонтальный и вертикальный контроль плиты. Для малоиспользуемых пандусов менее затратный метод (метод толкания) заключается в формировании и заливке пандуса тонким слоем песка или дробилки, дайте ему высохнуть, а затем толкните его в воду с помощью гусеничной машины. Если пандус заливают на берег, он должен быть примерно на том же уклоне, что и предполагаемый подводный уклон, чтобы предотвратить разрушение плиты при изменении уклона. Бетонные плиты, которые перемещаются на место, должны быть небольшими. Шестидюймовая бетонная плита шириной 12 футов и длиной 20 футов весит около девяти (9) тонн. Обычно плиты длиннее 20 футов необходимы для достижения соответствующей глубины. Шестидюймовая плита шириной 16 футов и длиной 30 футов весит около восемнадцати (18) тонн и обычно может толкаться бульдозером D-5, сохраняя при этом разумный контроль. Подъем и установка сборных железобетонных плит на подготовленное земляное полотно с помощью крана — это третий метод, с которым DGIF успешно справился.
Строительство пирса
После того, как сваи забиты, поперечные распорки и зажимы установлены, для строительства пирса требуются только общие столярные навыки. Тем не менее, следует проявлять большую осторожность, а также метод, используемый для забивки свай. Деревянные сваи никогда не следует использовать в условиях, когда сваи должны быть забиты через сплошную или трещиноватую породу или материал каменной наброски. Если деревянную сваю необходимо забить в почву с большим содержанием гравия, на кончике сваи следует использовать стальной башмак, чтобы защитить кончик сваи от «выметывания» во время забивания.
Сваи классифицируются как висячие или несущие. Висячие сваи развивают необходимую им прочность за счет трения, возникающего между внешней поверхностью сваи и окружающим грунтом. Несущие сваи забиваются на глубину, где они упираются в коренную породу и передают свои вертикальные нагрузки непосредственно в коренную породу. Длина сваи в проектах DGIF обычно определяется путем пробной забивки свай на площадке, чтобы выбрать различную длину сваи в зависимости от реальных условий подземных условий. Длина сваи также может быть определена в лаборатории инженером-геотехником по бурению грунта, взятому из шнека, установленного на барже; однако этот метод является дорогостоящим, и его следует использовать только тогда, когда он является наиболее рентабельным.
«Впрыскивание» свай в почву с содержанием ила или глины приводит к тому, что вода в окрестностях сильно замутняется взвешенными отложениями, что является экологически небезопасным и, как правило, является нарушением разрешений. Дальнейшее обсуждение будет ограничено забивкой свай. Существует несколько различных типов молотов, которые можно использовать для забивки свай. Для некоторых подземных условий подойдет любой тип молотка, но ни один молот не подходит для всех условий. Молоты для деревянных свай могут быть как гравитационными, так и силовыми (воздушные, паровые, дизельные, двухстороннего действия, одностороннего действия и т. д.). Как правило, гравитационный молот работает быстро в мягких грунтах, где можно использовать большой удар молота. В твердых грунтах необходимо использовать короткий удар гравитационного молота, чтобы предотвратить повреждение сваи; таким образом, время вождения увеличивается. Молоты одинарного и двойного действия используют пар или воздух и наносят больше ударов в минуту, чем гравитационный молот. Вибромолоты используют вибрации низкой или высокой частоты для ослабления трения и сцепления между грунтом и сваей, что позволяет свае проникнуть в почву. Вибрационные молоты обычно эффективны при работе с песком или мягким грунтом. Перед выбором типа забивки свай, длины или метода забивки необходимо проконсультироваться с инженером, специализирующимся на морском проектировании, или с морским подрядчиком.
Материалы
Любая конструкция хороша настолько, насколько хороши используемые материалы. Если был выбран хороший проект и методы строительства являются подходящими, надлежащие материалы и качество изготовления должны быть всем, что необходимо для обеспечения качественных лодочных аппарелей и пирсов. Надзор за качеством изготовления остается за владельцем или инженером. Следующие рекомендации могут служить руководством для выбора подходящих материалов.
Камень
При подготовке площадки для бетонирования поверх земляного полотна необходимо уложить не менее восьми дюймов уплотненного щебня (не круглого гравия). Камень VDOT № 5 или № 57 — хороший выбор. Если встречаются мягкие места или почву трудно уплотнить, плохой материал следует подрезать, а более крупный камень (диаметром от двух до четырех дюймов) следует использовать, чтобы обеспечить достаточную основу для меньшего камня. На рынке также есть геотекстильные ткани для «перекрытия» мягкого материала. Если какой-либо камень будет обнажаться по краям бетона, например, там, где бетон будет выше прилегающей земли, необходимо поместить под камень фильтрующую ткань хорошего качества и обернуть ее поверх камня. Ткань должна быть армирована каменной наброской для защиты фильтровальной ткани и, в конечном счете, камня под плитой. Если плита должна быть установлена на место, фильтрующая ткань не может быть установлена между бетоном и камнем.
Бетон
Бетон для пандусов, залитых на площадке, должен иметь давление не менее 3000 фунтов на квадратный дюйм (предпочтительно 4000 фунтов на квадратный дюйм), иметь 4-6% вовлечения воздуха, быть правильно обработанным, уложенным, укрепленным, выровненным и залитым так, чтобы заполнитель был примерно наполовину -дюйм ниже поверхности. DGIF успешно использовал бетонные пандусы, армированные арматурой номер четыре (диаметром 1/2 дюйма) марки 60, расположенной на расстоянии 12 дюймов от центра по длине и 18 дюймов по центру поперек. В пресной воде стальная арматура должна находиться на расстоянии 3″ от поверхности, дна и сторон бетона. Для этого требуется плита толщиной не менее шести дюймов. Для пандусов, с которых могут спускаться более крупные лодки (более 20 футов) или тяжелое оборудование, может подойти более толстый бетон, более крупная сталь и/или более прочный бетон. Поверхность бетона должна быть достаточно шероховатой, чтобы обеспечить хорошее сцепление даже с водорослями. Мы обнаружили, что садовые грабли с изогнутыми зубцами, изгибающимися от рукоятки, являются хорошим инструментом для проделывания канавок глубиной в четверть дюйма в свежем бетоне для сцепления. DGIF недавно получил V-образную канавку для улучшения сцепления. Эта отделка требует опытного бетонщика и не должна выполняться новичком.
Дерево и скобяные изделия
Древесина для пирсов должна быть обработана давлением, чтобы замедлить гниение. Исторически сложилось так, что древесина, обработанная CCA (хромированным арсенатом меди), была предпочтительной обработкой для строительства наших пирсов. Агентство по охране окружающей среды (EPA) сочло древесину, обработанную CCA, непригодной для использования в тех случаях, когда древесина может часто контактировать с кожей. Древесину, обработанную CCA, нельзя использовать в пресной воде. По этой причине мы используем обработанную CCA древесину, если древесина находится в постоянном контакте с соленой водой или в условиях постоянного увлажнения, например, сваи опор, поперечные связи, стрингеры палубы и балки. Для всех применений, которые могут вступить в контакт с кожей, мы решили использовать альтернативную обработку азолом меди (CA) для совместимости с оборудованием с гальваническим покрытием. Древесина CCA, которая будет находиться в постоянном контакте с соленой водой, должна быть обработана до уровня удерживания 2,5 фунта на кубический фут (pcf) древесины. Древесина, которая будет подвергаться брызгам соленой воды, должна иметь удержание 1,5 фунта на кубический фут.
