Вм 127м технические характеристики: Купить фрезерный станок ВМ 127М, цена, технические характеристики, фото. Продажа станка ВМ127М с доставкой по всей России и СНГ
Содержание
Вертикально фрезерный станок ВМ-127М (2003г.в.) КАП. РЕМОНТ 2011г.
Цена на 25.03.2023
Под заказ
Производитель:
Россия
ПОСЛЕ КАПИТАЛЬНОГО РЕМОНТА
Вертикально фрезерный станок ВМ-127М является аналогом станков 6Р13, 6Т13, FSS450R и предназначен для выполнения операций фрезерования различных деталей из черных и цветных металлов и их сплавов в условиях серийного и мелкосерийного производства.
Мощный привод главного движения и тщательно подобранные передаточные отношения обеспечивают оптимальные режимы обработки при различных условиях резания и полное использование возможностей режущего инструмента.
ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ СТАНКА ФРЕЗЕРНОГО ВМ-127М с МКИ
|
Характеристика |
ВМ-127М КАП. РЕМОНТ |
|---|---|
|
Размеры рабочей поверхности стола, мм |
400х1600 |
|
Наибольшее перемещение стола, мм:
|
1010 320 420 |
|
Наибольшее перемещение пиноли шпинделя, мм |
80 |
|
Угол поворота шпиндельной головки, град |
± 45 |
|
Расстояние, мм:
|
30-500 420 |
|
Количество скоростей шпинделя |
18 |
|
Частота вращения шпинделя,об/мин |
40.  ..2000 ..2000
|
|
Пределы подач, мм/мин:
|
25-1250 25-1250 8,3-416,6 |
|
Точность отсчета лимбов горизонтального и поперечного перемещений, мм |
0,05 |
|
Мощность электродвигателей, квт:
|
11 3 |
|
Масса ВМ-127М КАП. РЕМОНТ обрабатываемых деталей c приспособлением, кг |
450 |
|
Габариты, мм |
2680х2260х2500 |
|
Габариты упаковки(ДхШхВ), мм |
2830х2480(1340)х2625 |
|
Масса ВМ-127М КАП.  РЕМОНТ , кг РЕМОНТ , кг
|
4250 |
|
Масса ВМ-127М КАП. РЕМОНТ с упаковкой, кг |
4933 |
Станок фрезерный вм 127м технические характеристики
Фрезерный станок ВМ127: технические характеристики, схемы, эксплуатация
Известный в среде профессионалов фрезерный станок модели ВМ127 относится к давно забытой технике, распространенной еще в прошлом веке. Однако некоторые из его характеристик настолько уникальны, что до сих пор привлекают внимание специалистов. В связи с этим имеет смысл ознакомиться с основным функционалом и особенностями этого станка более подробно.
Сведения о производителе вертикально-фрезерного станка ВМ127
Производство станков этой серии было налажено Воткинским заводом еще в 1956 году. Само это предприятие было основано в далеком 1759 году, а сейчас является одним из ведущих производителей продукции широкого профиля.
Из его истории известно, что фрезерные станки марки ВМ127 являются прямыми аналогами других известных моделей, таких, например, как 6Р13, 6М13, а также ВМ127М и ВМ130.
Назначение станка
Станок вертикальный ВМ127 согласно ТУ предназначается для фрезерования деталей весом не более 450 кг (с оснасткой), изготавливаемых на основе чугуна, стали, а также ряда цветных металлов. Для их обработки применяются фрезы самого различного вида, а именно:
- Торцовый инструмент.
- Концевая и цилиндрическая оснастка.
- Радиусные и другие типы фрез.
На станке удается делать пазы произвольной ориентации, фрезеровать различные углы, рамки, а также зубчатые колеса.
Особенности его конструкции позволяют эффективно использовать возможности быстрорежущего инструмента. В случае перенастройки на полуавтоматический режим не исключено использование станка в составе группы аналогичных изделий.
Мощный привод ВМ127 и грамотно подобранные передаточные отношения в коробке передач (КС) создают идеальные условия для обработки различных исходных заготовок. Другой отличительной особенностью этого станка является простота обслуживания, а также возможность переналадки приспособлений и изменения инструментального набора. Наличие развитой системы смазки рабочих узлов обеспечивает надежное функционирование в критических условиях.
Технические характеристики
Универсальные характеристики изделия ВМ127 обеспечивают эффективную работу оборудования в различных пространственных плоскостях.
Габариты и масса станка
Основные пространственные характеристики приведены ниже:
- Полные размеры стола – 400х1600 мм.
- Его максимальное перемещение в продольном направлении – 1010 мм, в поперечном – 320 мм, а по вертикали – 420 мм.
- Максимум перемещения головки шпинделя – 80 мм.
При этом угол ее пространственного поворота составляет ± 45 градусов. Фиксированное расстояние от среза шпинделя станочного оборудования до поверхности стола составляет 30-500 мм.
Фиксированное расстояние от среза шпинделя станочного оборудования до поверхности стола составляет 30-500 мм.
Кроме того, для пользователя важны следующие характеристики:
- Число скоростей в коробке передач – 18.
- Частота оборотов шпинделя – 40-2000 в минуту.
- Предельные скорости: продольного движения – 25-1250 мм/мин (такое же значение имеет показатель поперечного смещения).
- Для вертикального перемещения оно равна 8,3-416,6 мм/мин.
- Погрешность отсчета перемещений по лимбам – 0,05 мм.
- Мощность встроенных электродвигателей: главной подачи – 11 кВт и привода подач – 3 кВт.
Также следует представить габариты станины в мм: 2680х2260х2500 и массу изделия, составляющую 4250 кг.
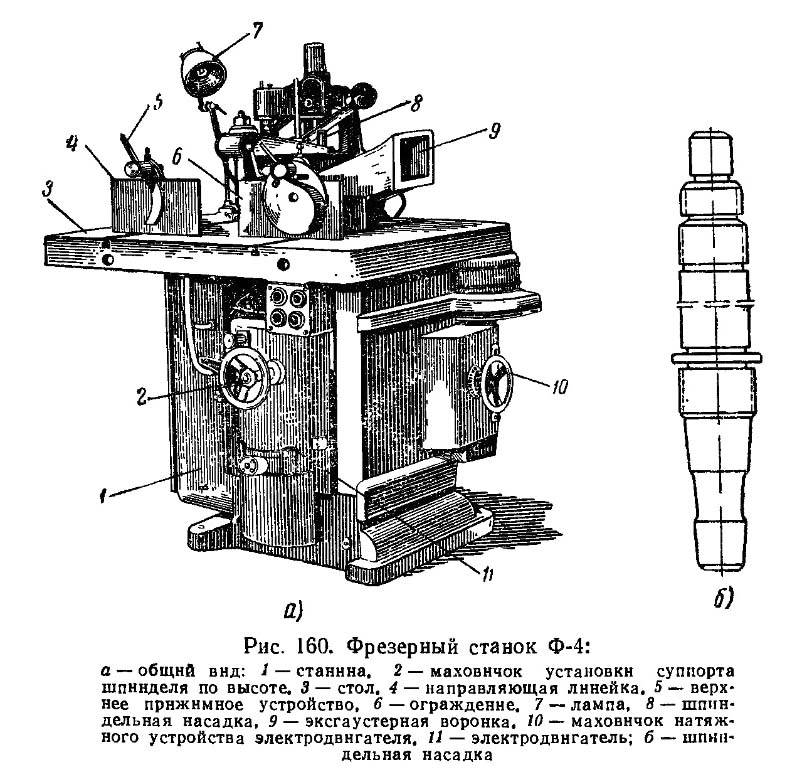
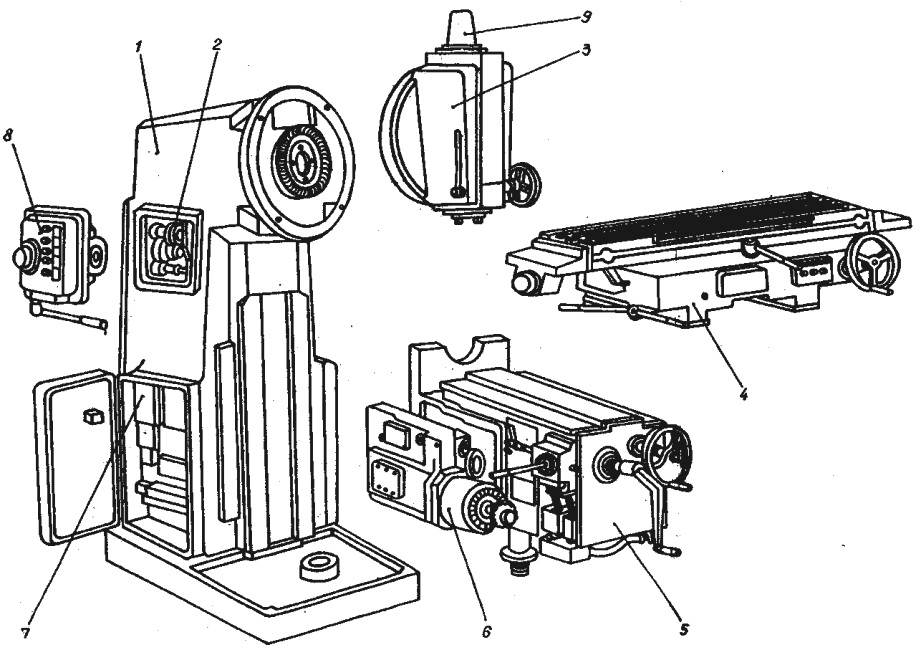
Общий вид вертикального фрезерного станка ВМ127
С тем, как выглядит внешний вид станка можно ознакомиться на фото слева. Из него следует, что данный образец состоит из трех основных частей, а именно:
- Вертикально расположенной станины с поворотной головкой.

- Инструментального стола с салазками и органами ручного управления.
- Рабочей консоли с подающим узлом.
Все эти узлы совмещены в едином сборном корпусе станка и обеспечивают его нормальное функционирование в различных режимах.
Расположение составляющих аппарата
Для ознакомления с расположением основных узлов достаточно исследовать приведенное ниже фото. Из него следует, что в составе ВМ127 имеются составляющие согласно приводимой далее спецификации.
Спецификация
Перечень оборудования станка представлен следующими позициями:
- Станина.
- Коробка скоростей (КС).
- Головка поворотная.
- Стол, оснащенный салазками.
- Рабочая консоль.
- Подающий узел.
- Система электрооборудования.
- КС шпинделя.
К числу составляющих ВМ127 следует отнести и механизм инструментального зажима.
Перечень органов управления фрезерным станком ВМ127
Для того чтобы представить, как расположены основные органы управления – достаточно ознакомиться с фото ниже.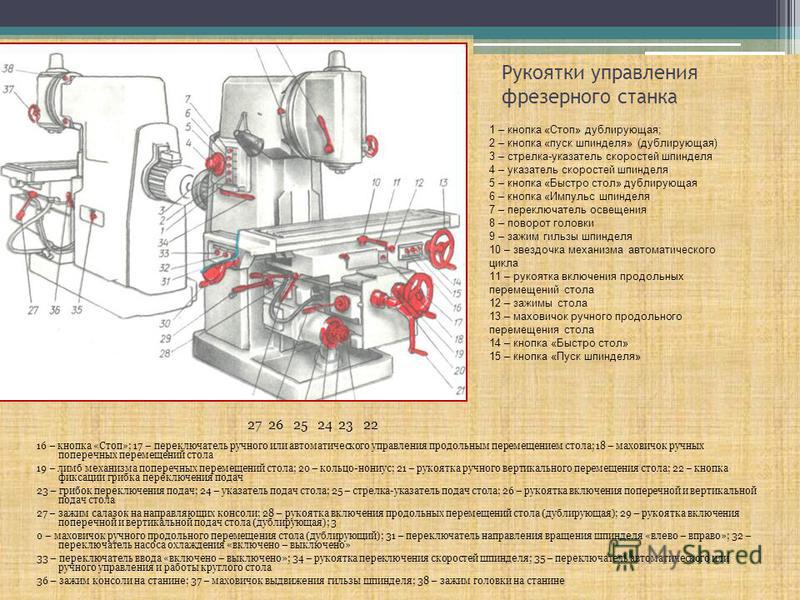
Расположение органов управления
Из приведенного фото видно, что основные органы управления расположились в удобных для этого зонах, а именно:
- на передней части стола;
- сбоку от поворотной головки;
- на боковинах станины.
Ручки управления подачей находятся прямо перед оператором чуть ниже уровня стола. К ним относятся:
- Маховики перемещений инструментального стола.
- Рукоятка его движения по вертикали.
- Рычаг включения продольного смещения.
- Переключатель режима управления.
На боковинах станины и сбоку от поворотной головки имеются следующие управляющие элементы:
- Кнопки включения и выключения станка и его основных рабочих узлов.
- Ручки поворота головки и зажимы гильзы.
- Грибок переключения подач и другие.
Помимо этого на передней панели рабочего стола и консоли располагаются и другие органы, включая различные зажимы и переключатели.
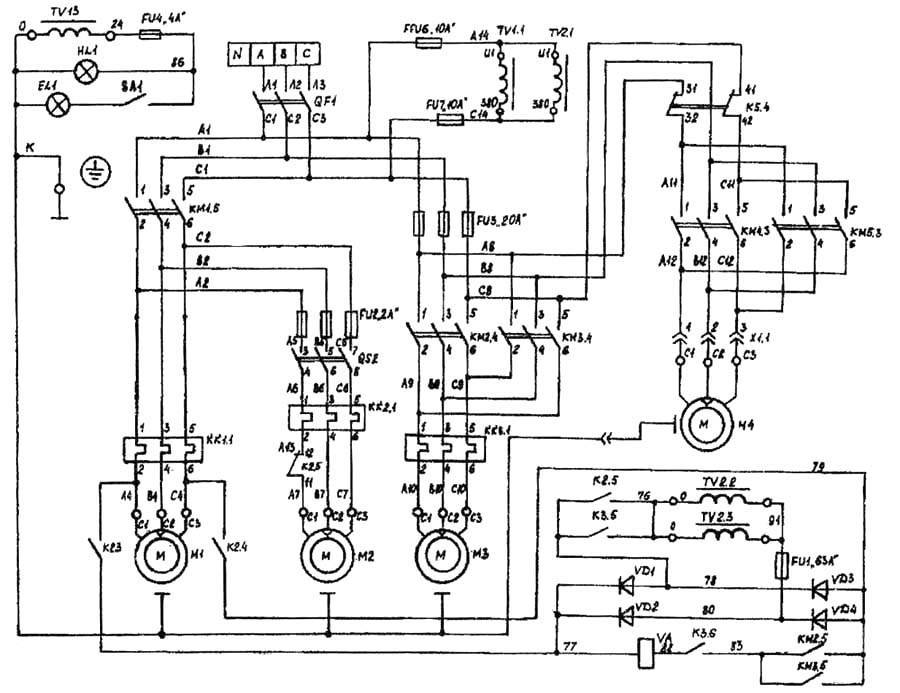
Электрическое оборудование
В электрооборудовании станка ВМ127 можно выделить следующие три части: силовая, управляющая и модуль питания.
Силовая часть
Эта составляющая электрической схемы представлена цепями питания основного и вспомогательных электродвигателей (М1, М2 и М3) и включает в себя следующие элементы:
- Контакторы пускателей.
- Предохранительные вставки.
- Тепловые реле.
- Элементы реверса.
Благодаря этим деталям обеспечивается работоспособность всех приводных систем и требуемая функциональность оборудования.
Управляющая часть
К управляющей части электрики относятся коммутирующие элементы (реле времени) с группой слаботочных контактов, переключающих режимы работы различных узлов. Сюда же входят защитные компоненты (предохранители и катушки индуктивности).
Обратите внимание! Схемы коммутации узлов станочного оборудования приводятся в таблицах.
Руководствуясь ими, можно будет выбрать требуемый режим работы.
Система питания
Питающие цепи электрооборудования обеспечивают:
- Преобразование переменного напряжения в постоянный потенциал, необходимый для питания реле.
- Трансформацию напряжения 220 Вольт до уровня, требуемого для осветительного прибора (лампочки).
Они включают в себя диодный мост, выключатели, а также предохранители и понижающий трансформатор.
Работа составных частей электрооборудования
Работа станка в наладочном режиме
Ниже рассматривается порядок функционирования различных узлов станочного агрегата под управлением, рассмотренного выше электрооборудования (в режиме наладки).
Зажим инструмента
Для зажима того или иного инструмента используется переключатель SA3, размещенный на боковом пульте (для этого его нужно перевести в положение «Зажим», одновременно удерживая рукой). В этом случае в работу вступает пускатель КМ4, подающий питание на двигатель функции зажима М4. Микрик SQ10 включает пускатель К5. 1, после чего тот устанавливается на самоблокировку и отключает М4, одновременно подготовив цепь для запуска мотора шпинделя.
1, после чего тот устанавливается на самоблокировку и отключает М4, одновременно подготовив цепь для запуска мотора шпинделя.
Включение, выключение и торможение шпинделя
Для запуска в работу шпинделя потребуется нажать SB7, после чего включаются пускатель под обозначением КМ1, а вслед за ним и реле КТ1 (КТ2). Через контакторы пускателя напряжение трехфазное 380 Вольт поступает на двигатель Ml, a KT2 своей нормально разомкнутой группой контактов осуществляет блокировку SB7.
Для того чтобы выключить шпиндель – достаточно нажать SB4, после чего напряжение снимается с КМ1, а также с КТ1, КТ2. Примерно через 1-2 секунды сработает пускатель К2, включающий торможение шпинделя. Еще через 5-6 секунд происходит его отключение (шпиндель полностью останавливается).
Включение насоса охлаждения
Для подачи питания на насос охлаждения предусмотрен переключатель QS2. Через его контакты напряжение 380 Вольт поступает на двигатель М2 (шпиндель в это время включен).
Аварийное выключение станка
В аварийных ситуациях следует нажать кнопку SB1 (SB2), посредством которой отключается цепь питания 110 Вольт. После этого она под действием пружины возвращается в исходное положение, обеспечивая возможность повторного включения оборудования.
Импульсное включение
Для того чтобы облегчить операцию смены скоростей шпинделя в станке предусмотрен импульсный режим включения электромотора M1 посредством кнопки SB5.
Электропривод подач
Для управления подачами предназначаются рукоятки, имеющие 3 фиксированные положения. В управляющие цепочки также входят выключатели SQ6, SQ8, используемые для продольного движения.
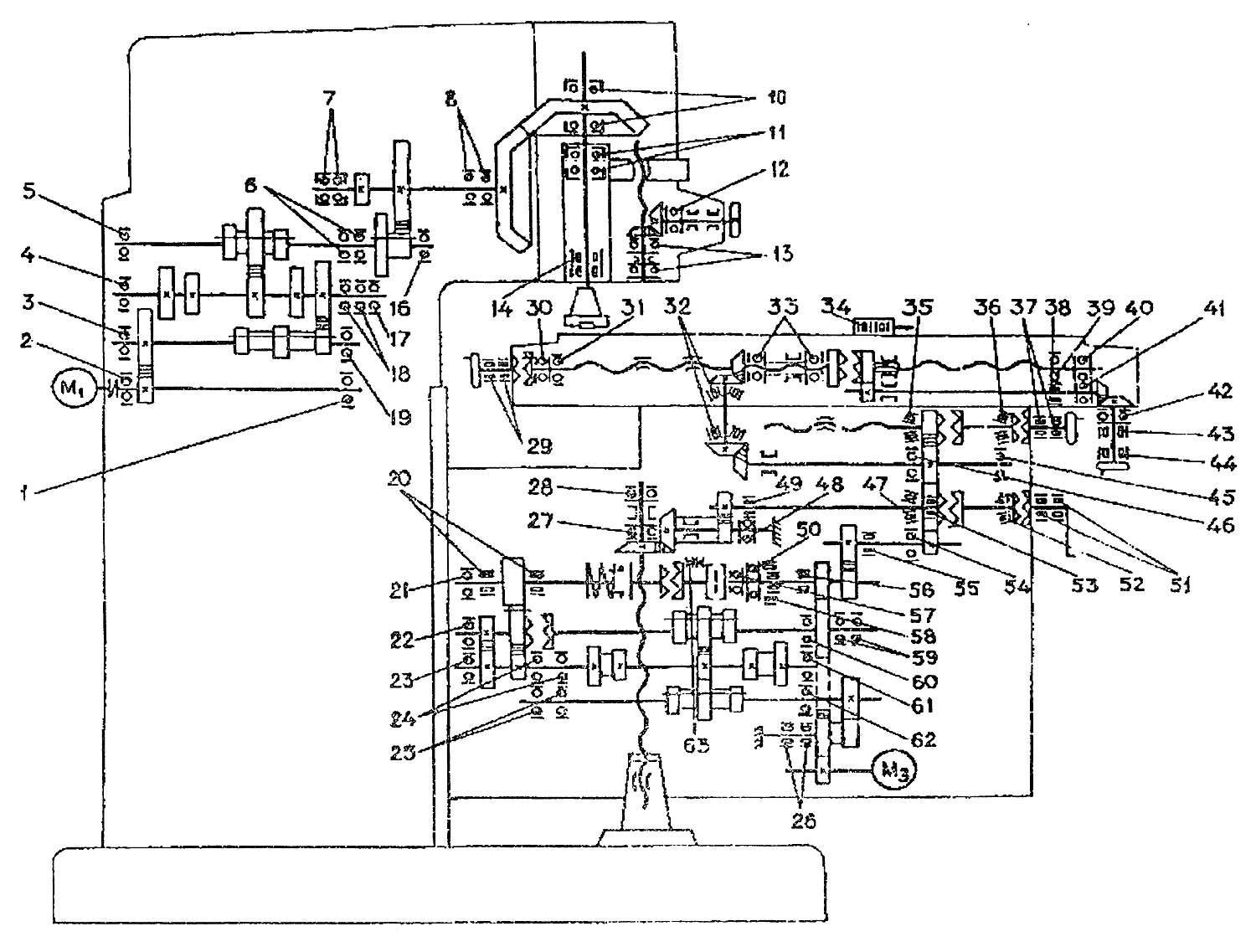
Кинематическая схема
Регулировка цепи торможения шпинделя
Процедура регулировки этой цепи необходима в случае отклонения временных параметров от нормы или по окончании ремонта КТ1 (КТ2). Для ее проведения потребуется:
- Запустить шпиндель.
- Спустя какое-то время выключить его.

- Одновременно с этим включить секундомер, а затем остановить его в момент щелчка, отчетливо слышимого при срабатывании пускателя К2.
- В том случае, если время задержки превышает 1 сек – провернуть регулятор КТ1 по часовой стрелке.
После этого следует повторно проверить задержку отключения шпинделя, добиваясь нужного момента включения. При его отклонении в другую сторону регулятор КТ1 нужно повернуть против хода часов. Настройка реле КТ2 аналогична.
Далее следует отключить кнопкой SB4 работающий шпиндель и одновременно с этим запустить секундомер, окончательно выключив его после полной остановки. Замеренное таким образом время не может превышать 6-ти секунд.
Принципиальные изменения конструкции станка ВМ127М после 2012 года
В 2012 году уральский завод освоил выпуск новой модели ВМ127М, которая претерпела следующие изменения:
- Конструкция расположенных вертикально направляющих была изменена. Вместо профиля типа «Ласточкин хвост» в ней стала применяться П-образная форма, что сделало возможным обрабатывать детали весом до 800 кг.

- Механическая коробка станка заменена серводвигателем, обеспечивающим бесступенчатое переключение подач.
- В моделях с цифровой индикацией данных обработки импортные комплектующие изделия заменены отечественными.
В заключение отметим, что новые модели ВМ127М способны облегчить процесс обработки деталей, одновременно повышая производительность операций.
Votkinski Zavod BM127M Вертикальный станок
Этот сайт использует куки. Продолжая просматривать Exapro, вы соглашаетесь использовать куки на нашем сайте.
Меню
- Home
- Купить
- Электроника 108
- Машины для приготовления пищи 3448
- Металл — литейное производство / прессы 1497
- Металл — станки 10307
- Металл — листовой металл 4441
- Другое промышленное оборудование 2527
- Упаковка — кондиционирование 3881
- Фармацевтическая — Химическая 2169
- Пластик — резина 2648
- Печатные машины
.
Вертикальный фрезерный станок4h 5h Фрезерный станок револьверный фрезерный станок
0 штук выбрано, всего
Посмотреть детали
- Стоимость доставки:
- Зависит от количества заказа.
- Время выполнения:
- 30 дней после получения оплаты
- Персонализация:
Индивидуальный логотип (Мин.Заказ: 1 комплект)
Индивидуальная упаковка (Мин. Заказ: 1 комплект)
Подробнее
Настройка графики (Мин.Заказ: 1 комплект) Меньше
Образцы: 3 170,00 $ / комплект, 1 комплект (минимальный заказ): Купить образцы ,
New Ce, Iso Вертикально-расточной станок с цилиндрической головкой Bm200
NEW CE, ISO Сверлильно-фрезерный станок с цилиндрической головкой ISO BM200
Вертикально-фрезерно-расточный станок BM200 сочетает в себе все функции и простую работу традиционной сверлильно-фрезерной машины с цилиндрической головкой и простой в эксплуатации Система управления SJMC EasyTouch. Благодаря совместной работе стандартного механического маховика и электронного маховика легко может быть выполнена расточка корпуса цилиндра и обработка плоскости фрезерования.С превосходной системой EasyTouch даже новая рука может легко научиться управлять машиной.
Благодаря совместной работе стандартного механического маховика и электронного маховика легко может быть выполнена расточка корпуса цилиндра и обработка плоскости фрезерования.С превосходной системой EasyTouch даже новая рука может легко научиться управлять машиной.
BM200 особенно подходит для точной работы переоборудованного двигателя: расточка, фрезерование, обработка ступенчатых отверстий, обработка канавок ….. гарантирует высокую стабильность всей обработки
Особенность:
1. шпиндель с расточным резцом может направленно запускать и останавливать расточку
2.Точное управление нулевым набором оси Z
3. Точность повторного позиционирования оси Z ≤ 0,005 мм
4. QuickChange — шпиндель
5. Одностороннее включение расточки 1,2 мм
6. Дополнительный 3-осевой УЦИ.
Характеристика:
1. Операционная система EasyTouch
Операционная система EasyTouch
Левое и правое движение стола, подъем и опускание шпинделя, а также вращение Все шпиндели приводятся в движение серводвигателем, который, комбинируя со специальной системой EasyTouch, обеспечивает точное числовое управление движением трех осей на всей машине.
2. Малый размер для выдачи высокого крутящего момента
BM160 имеет компактную конструкцию, с рабочей площадью 1,5 м 2 меньше, чем у аналогичных машин. Тем не менее, с максимальным рабочим моментом 120 Нм, его небольшой размер демонстрирует мощные возможности обработки. Благодаря шпинделю BM160, приводимому в действие серводвигателем мощностью 4 кВт и оснащенному системой ременной передачи тяжелой нагрузки, шпиндель способен поддерживать постоянный мощный выходной крутящий момент 120 Нм при 200–500 об / мин и даже поддерживать постоянную мощность 100 Н.М на более высокой скорости от 500 до 780 об / мин. Высокий крутящий момент на высокой скорости обеспечивает высокую эффективность BM200.
Высокий крутящий момент на высокой скорости обеспечивает высокую эффективность BM200.
3. Точная и мощная система шпинделей
Продуманно спроектированная система шпинделей с 3 шпинделями от малого до большого, вы можете легко удерживать один из них одной рукой, плюс более хитрая конструкция с быстрой сменой, что способствует на смену шпинделя в течение 5 минут человеком. Несмотря на то, что все шпиндели изящны, их режущая способность очень высока.Проникновение режущего инструмента с одной стороны шпинделя MA52 и MA60 может достигать 1,5 мм с точностью сверления 0,005 мм.
4. Опциональная фрезерная головка MP300 оснащена 8 режущими кромками, , а подача фрезерования за один раз для облицовки составляет 1 мм, так что, независимо от чугунной или алюминиевой детали, может быть достигнута идеальная шероховатость поверхности. С плоскостностью, превышающей 0,013 мм / 300 мм, его точность обработки обеспечивает эффект заземления и, таким образом, он может заменить заземление фрезерной обработкой.
Основные технические характеристики
Модель | BM200 | 9012 9012 901 9015 (901) (901) 9000 (5) (929) (929) (929) (9000) (9000) (9000) | Без | |
Буровая емкость | 200 мм | |||
Макс.Глубина сверления | 350 мм | |||
Макс. Зона фрезерования (Д × Ш) | 300 × 920 мм | Без фрезерования | ||
Частота вращения шпинделя | 50 ~ 1000 об / мин Бесступенчато | |||
| 9015 9105 9015 9005 100000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000009000 10 ~ 900 мм / мин (0,01 ~ 0,2 мм / об) | ||||
Номинальный крутящий момент расточки при 50-500 об / мин | 58 Н. | |||
Номинальный крутящий момент расточки при 500-780 об / мин | 22 NM | |||
Траверса шпинделя | 600 мм | |||
0 ~ 700 мм | ||||
Расстояние между осью шпинделя и вертикальной плоскостью каретки | 380 мм | |||
Продольная подача рабочего стола по электрике | 9 мин. | Ручное движение | ||
Быстрый продольный рабочий стол с электрическим приводом | 1200 мм / мин | Ручное движение | ||
0000 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 9 5 9 5 000 000 5 5 5 5 5 5 000 000 000 000 9 000 000 000 000 9 000 5 000 000 000 000 000 000 000 Рабочий стол00 000 000 00000 | ||||
Крестовина рабочего стола | 100 мм | |||
Рабочий стол Размер (Д × Ш) | 400 × 1350 мм | |||
скучный точность | Н7 | |||
округлость для расточки | 0. | |||
Рабочей точность | Цилиндричности для расточных | 0,015 мм / 300 мм | ||
плоскостности для фрезерования | 0,0127 мм / 305 мм | Без фрезерования | ||
Шероховатость поверхности | Расточка | Ra 0,8 | ||
Фрезерование | Ra 0.8 | без фрезерования | ||
Мощность двигателя шпинделя | 2 кВт | |||
Мощность двигателя рабочего стола | 0,401 9015 0,401 9355 935 Руда 0,015 9355 935 | |||
Мощность двигателя шпинделя по вертикали | 0,4 кВт | |||
Габаритные размеры (Д × Ш × В) | 1870 × 1600 × 2000 мм | |||
Размер упаковки | 2200 × 1450 × 2250 мм | |||
Н. | 1900 кг | |||
G.W. | 2300 кг | |||
 M
M 005 мм
005 мм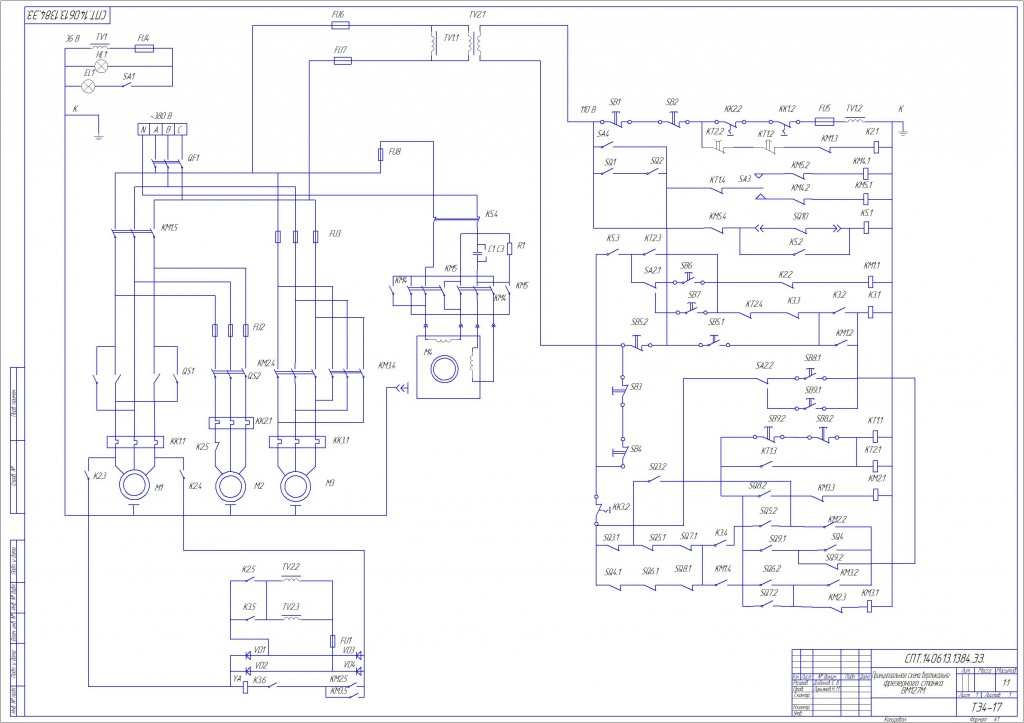 W.
W.Реальные фотографии:
Аксессуар:
Стандартная экспортная упаковка:
,
63-узловой кластер EKS, работающий на одном экземпляре с Firecracker
Этот пост является частью серии постов, посвященных автоматизации Firecracker . В настоящее время он состоит из
В настоящее время он состоит из
следующие сообщения:
- Часть I:
Автоматизация запуска виртуальных машин на основе ванильных облачных образов на Firecracker - Часть II (текущая)
Разрабатывая исходный код, созданный для автоматизации создания виртуальных машин Firecracker, этот пост покажет, как мы можем
создать кластер, совместимый с EKS (как в Amazon EKS), с несколькими узлами, где каждый
узел будет виртуальной машиной, работающей на одном хосте, благодаря Firecracker .
Весь исходный код доступен в
этот репозиторий GitLab.
Создание кластера EKS из виртуальных машин Firecracker
EKS? Разве это не управляемый AWS сервис Kubernetes? Это, конечно, так, но это также недавно объявленный открытый исходный код и
предоставлен Amazon Web Services. Последний
более точно называется EKS Distro (EKS-D), а его исходный код доступен на
этот репозиторий Github.
В частности, будет использоваться совместимый с EKS-D дистрибутив Ubuntu на основе оснастки, поскольку он
довольно прост в установке и эксплуатации на виртуальных машинах Ubuntu. Интересно, что это распределение основано на
Интересно, что это распределение основано на
microk8s, и помечен как « EKS-совместимый Kubernetes ». « Совместимость » в этом контексте
похоже, что это своего рода microk8s, «одетый как» EKS, поскольку он включает в себя бинарный файл eks , который работает, как и ожидалось.
в другом месте. Но поскольку он основан на microk8s, то, наверное, подходит только для тех случаев, когда есть microk8s.
С помощью кода, разработанного в рамках предыдущего поста, можно создать настраиваемое количество виртуальных машин Firecracker.
автоматически, с SSH без пароля, настроенным с пользователем fc , который также может работать без пароля sudo . Учитывая это и
что нам нужно установить и настроить экземпляры для установки EKS-D Ubuntu, Ansible был
идеальный кандидат для этой автоматизации.
Весь код Ansible можно найти в
доступный /
папка. Я не являюсь экспертом по Ansible, поэтому код подлежит множеству улучшений (не стесняйтесь присылать MR!). Код должен быть
Код должен быть
совершенно не требует пояснений — за исключением части инвентаря, которая потребовала некоторых дополнительных сценариев оболочки, которые можно найти
в
05-install_eks_via_ansible.sh
файл.
Есть две проблемы, которые нам нужно решить с помощью сценария оболочки:
Создать динамическую инвентаризацию. По сути, количество виртуальных машин основано на переменной
NUMBER_VMS, определенной в
переменныхфайл. Первый узел будет считаться ведущим EKS, а все остальные узлы рабочими.После установки оснастки
екскаждая нода будет отдельным «мастером». Для формирования единого кластера нам нужно запустить 9Команда 0027 eks add-node в мастере для каждого узла и эквивалентная командаeks joinв узлах. Обе команды используют токен,
который может быть сгенерирован автоматически (не удобно для этого варианта использования) или предоставлен. Сценарий оболочки генерирует эти токены
и предоставить их как переменные: токен для каждого узла и список всех токенов для главного узла.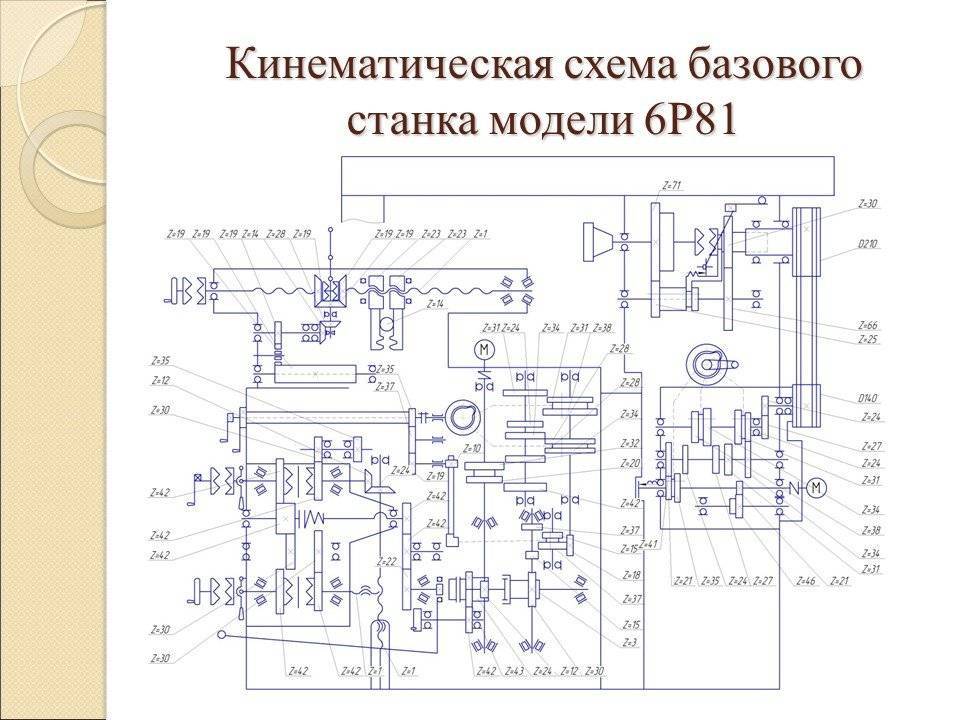
Вы можете запустить этот код в своей среде. Обзор
README.md
файл и убедитесь, что firecracker установлен и правильно настроен, как описано в
первый пост. 4 ГБ и 2
ядер кажется разумным минимумом ресурсов, необходимых для каждой виртуальной машины для запуска успешного кластера, совместимого с EKS-D. Регулировать
переменных в вашу среду.
Создание кластера EKS с 63 узлами на экземпляре r5d.metal
Не было бы здорово создать большой кластер EKS, работающий на множестве ВМ… и все это на одном «большом» хосте? Это
эксперимент я провожу. Я выбрал экземпляр r5d.metal (96 ядер, 768 ГБ ОЗУ,
Локальный твердотельный накопитель NVMe емкостью 3,6 ТБ) как лучшее сочетание большого количества ядер, оперативной памяти и локального хранилища (поскольку мы будем создавать
«тома» для виртуальных машин, и это должно быть максимально быстро). Затем запустил 100 виртуальных машин Firecracker и создал EKS.
кластер…
Я установил
количество вилок Ansible
до 32, чтобы иметь больше параллелизма. Однако похоже даже при низком параллелизме есть ошибки с экс джоин
операция вызвана ограничениями на максимальное количество запросов к kubeapi-серверу, поэтому
эта часть кода Ansible сериализуется.
Виртуальные машины с 2 ядрами и 4 ГБ ОЗУ. Код сработал. Скрипты, ансибл, все заработало. Чуть позже, через 4 часа,
Кластер EKS из 100 узлов, созданный в одном экземпляре, работал:
Однако нагрузка превышала 200 , и хотя использование памяти было приемлемым (2/3 оперативной памяти системы), кластер был
непригодный для использования. Но что более важно, я обнаружил интересный факт: kubectl get узлов вернули бы только 63 узла, несмотря на
99 операций соединения узлов были возвращены как успешные. Я особо не копал, но тут видимо есть ограничение
(внутреннее ограничение microk8s? В любом случае не уверен, что microk8s был тщательно протестирован с таким большим количеством кластеров
экземпляры…).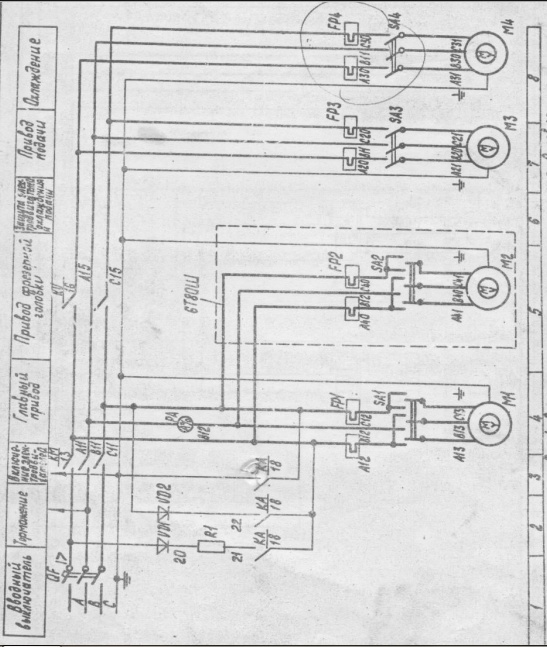
Поэтому я решил повторить операцию, но с «всего» 63 ВМ (ведь kubectl get узлов больше не вернут…) и
увеличьте характеристики ВМ до 4-ядерных (виртуальная машина главного узла также была загружена более чем на 100%) и 16 ГБ ОЗУ (16 ГБ * 63 — это
больше, чем доступная оперативная память системы, но Firecracker допускает чрезмерную нагрузку как на память, так и на ЦП, если не все
процессы используют всю выделенную оперативную память). Это сработало лучше, кластер был успешно создан и kubectl получили узлы
сообщил на этот раз все узлы. Все примерно за 3 часа, сейчас при приемлемой загрузке процессора:
ubuntu@ip-172-31-93-220:~$ kubectl получить узлы ИМЯ СТАТУС РОЛИ ВОЗРАСТ ВЕРСИЯ id1394205090 Готов <нет> 116м v1.18.9-екс-1-18-1 id0890521735 Готов <нет> 93м v1.18.9-екс-1-18-1 id1865400204 Готов <нет> 85м v1.18.9-екс-1-18-1 id3176525421 Готов <нет> 79м v1.18.9-екс-1-18-1 id0289819564 Готов <нет> 97м v1.18.9-екс-1-18-1 id2364814864 Готов <нет> 101м v1.18.9-экс-1-18-1 id1576708969 Готов <нет> 62м v1.18.9-екс-1-18-1 id0839217590 Готов <нет> 95м v1.18.9-eks-1-18-1 id2385921825 Готов <нет> 122м v1.18.9-екс-1-18-1 id2162009048 Готов <нет> 99м v1.18.9-екс-1-18-1 id0032931251 Готов <нет> 94м v1.18.9-екс-1-18-1 id1397218544 Готов <нет> 83м v1.18.9-екс-1-18-1 id2509806641 Готов <нет> 72м v1.18.9-екс-1-18-1 id1391026381 Готов <нет> 56м v1.18.9-экс-1-18-1 id3180021860 Готов <нет> 132м v1.18.9-екс-1-18-1 id2766207659 Готов <нет> 130м v1.18.9-eks-1-18-1 id2417208994 Готов <нет> 67м v1.18.9-екс-1-18-1 id0037315342 Готов <нет> 126м v1.18.9-екс-1-18-1 id2870908982 Готов <нет> 124м v1.18.9-екс-1-18-1 id0303528979 Готов <нет> 96м v1.18.9-екс-1-18-1 id2443620467 Готов <нет> 100м v1.18.9-eks-1-18-1 id2671116621 Готов <нет> 89м v1.18.9-экс-1-18-1 id1153919941 Готов <нет> 127м v1.18.9-екс-1-18-1 id1618117867 Готов <нет> 114м v1.18.9-екс-1-18-1 id1345427105 Готов <нет> 128м v1.18.9-екс-1-18-1 id1578112374 Готов <нет> 133м v1.18.9-екс-1-18-1 id2260431270 Готов <нет> 134м v1.
18.9-екс-1-18-1 id1192720052 Готов <нет> 131м v1.18.9-екс-1-18-1 id0009731256 Готов <нет> 86м v1.18.9-екс-1-18-1 id0772428726 Готов <нет> 112м v1.18.9-экс-1-18-1 id2345631371 Готов <нет> 115м v1.18.9-екс-1-18-1 id0866401320 Готов <нет> 90м v1.18.9-eks-1-18-1 id2295817569 Готов <нет> 121м v1.18.9-екс-1-18-1 id3195221619 Готов <нет> 105м v1.18.9-екс-1-18-1 id3212514411 Готов <нет> 64м v1.18.9-екс-1-18-1 id2530228284 Готов <нет> 58м v1.18.9-екс-1-18-1 id1582731567 Готов <нет> 125м v1.18.9-екс-1-18-1 id0234531685 Готово <нет> 104 м v1.18.9-экс-1-18-1 id1170407709 Готов <нет> 129м v1.18.9-екс-1-18-1 id2377401525 Готов <нет> 129м v1.18.9-екс-1-18-1 id2782930529 Готов <нет> 109м v1.18.9-екс-1-18-1 id2316206444 Готов <нет> 88м v1.18.9-екс-1-18-1 id0886017938 Готов <нет> 120м v1.18.9-eks-1-18-1 id2886224137 Готов <нет> 135м v1.18.9-екс-1-18-1 id2137806005 Готов <нет> 74м v1.18.9-екс-1-18-1 id3252717927 Готов <нет> 136м v1.18.9-экс-1-18-1 id1421103858 Готов <нет> 111м v1.18.9-екс-1-18-1 id2048704797 Готов <нет> 102м v1.
18.9-екс-1-18-1 id1892014011 Готов <нет> 106м v1.18.9-екс-1-18-1 id0719403446 Готов <нет> 61м v1.18.9-екс-1-18-1 id1134505657 Готов <нет> 82м v1.18.9-екс-1-18-1 id0023523299 Готов <нет> 118м v1.18.9-екс-1-18-1 id2477116066 Готов <нет> 81м v1.18.9-екс-1-18-1 id0469627718 Готов <нет> 77м v1.18.9-экс-1-18-1 id1020429150 Готов <нет> 113м v1.18.9-екс-1-18-1 id1471722684 Готов <нет> 91м v1.18.9-екс-1-18-1 id1417322458 Готов <нет> 75м v1.18.9-eks-1-18-1 id0107720352 Готов <нет> 69м v1.18.9-екс-1-18-1 id2957917156 Готов <нет> 108м v1.18.9-екс-1-18-1 id0573606644 Готов <нет> 3ч54м v1.18.9-экс-1-18-1 id1377114631 Готов <нет> 117м v1.18.9-екс-1-18-1 id0468822693 Готов <нет> 119м v1.18.9-экс-1-18-1 id0660532190 Готов <нет> 71м v1.18.9-екс-1-18-1
Несмотря на все это, кластер нельзя было использовать. Большинство операций выполнялись долго, а некоторые приводили к ошибкам.
Вероятно, microk8s, на котором основан этот EKS-D, не предназначен для этих целей 😉
Заключительные слова
Эксперимент с 63 и 100 узлами был скорее забавным упражнением и проверкой скриптов и кода Ansible. Однако,
Однако,
код представлен
весьма полезен, особенно для тестирования сценариев. Я могу создать на своем ноутбуке 3-узловой кластер EKS (2 ядра, 4 ГБ ОЗУ на
узел) менее чем за 5 минут, и все это с помощью однострочной команды. И уничтожь все это за секунды еще одной строчкой . Этот
позволяет быстро тестировать EKS-совместимые кластеры Kubernetes. Все благодаря Firecracker и EKS-D, оба с открытым исходным кодом
компоненты, выпущенные AWS. Спасибо!
Интерактомный анализ последовательного элюирования ментальной бомбы 1 Убиквитинлигаза раскрывает новую роль в росте дендритных шипов
1.
Ито М., Ким С.-Х., Паларди Г., Ода Т., Цзян Ю.-Дж., Мауст Д., Йео С.-Ю., Лорик К., Райт Г.Дж., Ариза-Макнотон Л., Вайсман А. М., Льюис Дж., Чандрасехараппа С. К., Читнис А. Б. (2003) Ментальная бомба представляет собой убиквитинлигазу, которая необходима для эффективной активации передачи сигналов Notch с помощью Delta. Дев. Клетка
4, 67–82 [PubMed] [Google Scholar]
2.
Ван Иден Ф.Дж., Гранато М., Шах У., Бранд М., Фурутани-Сейки М., Хафтер П., Хаммершмидт М., Гейзенберг С.П., Цзян Ю.Дж., Кейн Д.А., Келш Р.Н., Маллинс М.К., Одентал Дж., Варга RM, Allende ML, Weinberg E.S., Nüsslein-Volhard C. (1996) Мутации, влияющие на формирование и формирование сомитов у рыбок данио, Danio rerio. Разработка
123, 153–164 [PubMed] [Google Scholar]
3.
Хэддон С., Цзян Ю. Дж., Смитерс Л., Льюис Дж. (1998) Передача сигналов Delta-Notch и формирование паттернов дифференцировки сенсорных клеток в ухе рыбок данио: данные мутанта с ментальной бомбой. Разработка
125, 4637–4644 [PubMed] [Google Scholar]
4.
Лоусон Н.Д., Шеер Н., Фам В.Н., Ким С.Х., Читнис А.Б., Кампос-Ортега Дж.А., Вайнштейн Б.М. (2001)Передача сигналов Notch необходима для артериально-венозной дифференцировки во время эмбрионального развития сосудов. Разработка
128, 3675–3683 [PubMed] [Google Scholar]
5.
Capoccia BJ, Jin R.U., Kong Y., Peek RM, Jr., Fassan M., Rugge M., Mills JC (2013)Убиквитинлигаза Mindbomb 1 координирует созревание секреторных клеток желудочно-кишечного тракта. Дж. Клин. Вкладывать деньги. 123, 23–27 [бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Клин. Вкладывать деньги. 123, 23–27 [бесплатная статья PMC] [PubMed] [Google Scholar]
6.
Ку Б.-К., Юн М.-Дж., Юн К.-Дж., Им С.-К., Ким Ю.-Ю., Ким Ч.-Х., Су П.-Г., Ян Ю. Н., Конг Ю.-Ю. (2007) Обязательная роль ментальной бомбы-1 в передаче сигналов развития млекопитающих. PLoS Один
2, е1221. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7.
Лю Л.-Дж., Лю Т.-Т., Ран Ю., Ли Ю., Чжан X.-Д., Шу Х.-Б., Ван Ю.-Ю. (2012) Убиквитинлигаза E3 MIB1 негативно регулирует базальный уровень IκBα и модулирует активацию NF-κB. Сотовый рез. 22, 603–606 [бесплатная статья PMC] [PubMed] [Google Scholar]
8.
Альтшул С.Ф., Мэдден Т.Л., Шеффер А.А., Чжан Дж., Чжан З., Миллер В., Липман Д.Дж. (1997) Gapped BLAST и PSI-BLAST: новое поколение программ поиска белковых баз данных. Нуклеиновые Кислоты Res. 25, 3389–3402 [бесплатная статья PMC] [PubMed] [Google Scholar]
9.
Takeuchi T., Heng HH, Ye CJ, Liang S.-B., Iwata J., Sonobe H., Ohtsuki Y. (2003) Понижающая регуляция новой молекулы, связывающей актин, скелетотрофина, при злокачественной меланоме.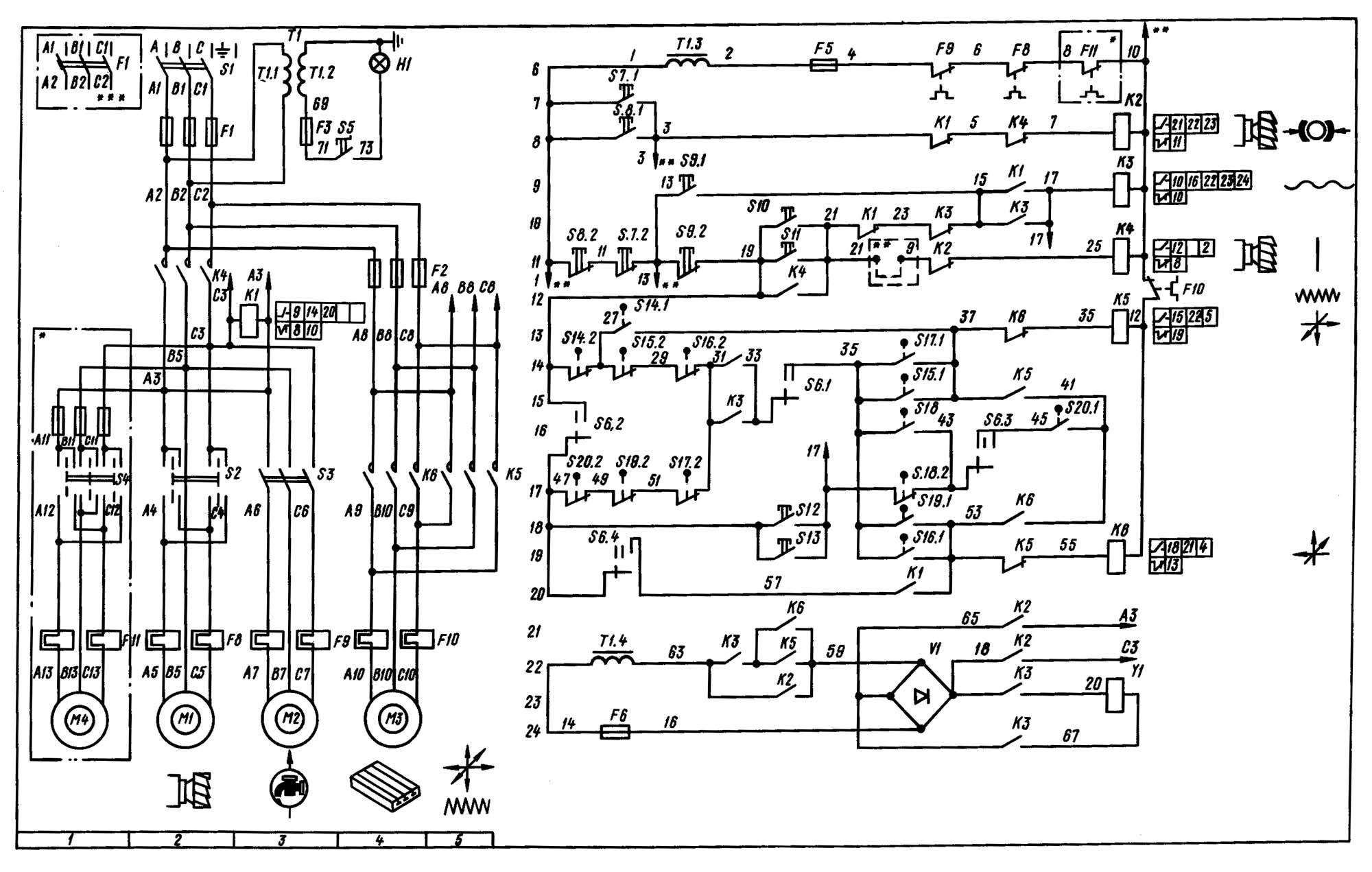 Являюсь. Дж. Патол. 163, 1395–1404 [бесплатная статья PMC] [PubMed] [Google Scholar]
Являюсь. Дж. Патол. 163, 1395–1404 [бесплатная статья PMC] [PubMed] [Google Scholar]
10.
Ку Б.-К., Лим Х.-С., Сон Р., Юн М.-Дж., Юн К.-Дж., Мун Дж.-С., Ким Ю.-В., Квон М.- C., Yoo K.-W., Kong M.-P., Lee J., Chitnis A.B., Kim C.-H., Kong Y.-Y. (2005) Бомба разума 1 необходима для создания функциональных лигандов Notch для активации Notch. Разработка
132, 3459–3470 [PubMed] [Google Scholar]
11.
Юн К.-Дж., Ли Х.-Р., Джо Ю.С., Ан К., Юнг С.-Ю., Чон М.-В., Квон С.-К., Ким Н.-С., Чон Х.-В., Ан С.-Х., Ким К.-Т., Ли К., Ким Э., Ким Дж.-Х., Чой Дж.-С., Каанг Б.-К., Конг Ю.-Ю. (2012) Бомба разума-1 является важным модулятором долговременной памяти и синаптической пластичности через сигнальный путь Notch. Мол. Мозг
5, 40. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12.
Чо Э.-А., Ляо Л., Чжоу Дж.-Ю., Ченг Д., Дуонг Д.М., Джин П., Цай Л.-Х., Пэн Дж. (2007) Морфогенез нейронов регулируется взаимодействием между циклин-зависимая киназа 5 и убиквитинлигаза интеллектуальная бомба 1.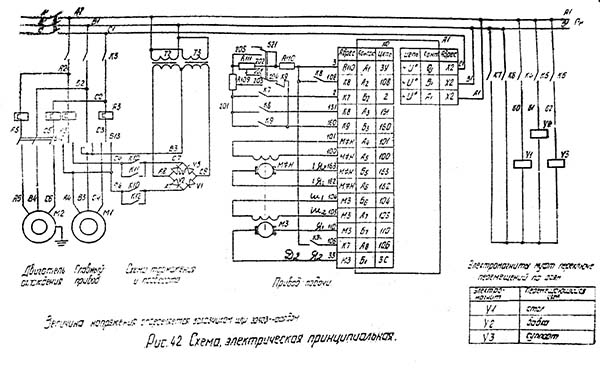 J. Neurosci. 27, 9503–9512 [бесплатная статья PMC] [PubMed] [Google Scholar]
J. Neurosci. 27, 9503–9512 [бесплатная статья PMC] [PubMed] [Google Scholar]
13.
Тессье-Лавин М., Гудман К.С. (1996) Молекулярная биология управления аксонами. Наука
274, 1123–1133 [PubMed] [Google Scholar]
14.
Cline HT (2001)Развитие дендритной ветви и синаптогенез. Курс. мнение Нейробиол. 11, 118–126 [PubMed] [Google Scholar]
15.
Нимчинский Э. а, Сабатини Б. Л., Свобода К. (2002) Структура и функция дендритных шипиков. Анну. Преподобный Физиол. 64, 313–353 [PubMed] [Google Scholar]
16.
Scheiffele P. (2003) Передача сигналов между клетками во время образования синапсов в ЦНС. Анну. Преподобный Нейроски. 26, 485–508 [PubMed] [Google Scholar]
17.
Квон Д.Ю., Димитриади М., Терзич Б., Кейбл С., Харт А.С., Читнис А., Фишбек К.Х., Бернетт Б.Г. (2013)Убиквитинлигаза Е3 бомба разума 1 убиквитинирует и способствует деградации выживания белка двигательного нейрона. Мол. биол. Клетка
24, 1863–1871 [бесплатная статья PMC] [PubMed] [Google Scholar]
18.
Berndt JD, Aoyagi A., Yang P., Anastas JN, Tang L., Moon R.T. (2011) Mindbomb 1, убиквитинлигаза E3, образует комплекс с RYK для активации передачи сигналов Wnt/{beta}-catenin. Дж. Клеточная биология. 194, 737–750 [бесплатная статья PMC] [PubMed] [Google Scholar]
19.
Ценг Л.-К., Чжан С., Ченг С.-М., Сюй Х., Хсу С.-Х., Цзян Ю.-Дж. (2014) Новые классы белков, взаимодействующих с мозговыми бомбами, идентифицированы с помощью двухгибридных экранов дрожжей. PLoS Один
9, e93394. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20.
Манн М., Кулак Н.А., Нагарадж Н., Кокс Дж. (2013) Грядущий век полных, точных и вездесущих протеомов. Мол. Клетка
49, 583–590 [PubMed] [Google Scholar]
21.
Чжан Ю., Фонслоу Б.Р., Шан Б., Бэк М., Йейтс Дж. Р. (2013) Анализ белков с помощью Shotgun / Bottom-up Proteomics. хим. Rev. 113, 2343–2394 [бесплатная статья PMC] [PubMed] [Google Scholar]
22.
Гстайгер М., Эберсолд Р. (2009) Применение протеомики на основе масс-спектрометрии к генетике, геномике и сетевой биологии. Нац. Преподобный Жене. 10, 617–627 [PubMed] [Google Scholar]
Нац. Преподобный Жене. 10, 617–627 [PubMed] [Google Scholar]
23.
Несвижский А.И. (2012) Вычислительные и информационные стратегии для идентификации специфических партнеров взаимодействия белков в экспериментах по масс-спектрометрии аффинной очистки. протеомика
12, 1639–1655 [бесплатная статья PMC] [PubMed] [Google Scholar]
24.
Риго Г., Шевченко А., Рутц Б., Вильм М., Манн М., Серафин Б. (1999) Общий метод очистки белков для характеристики белковых комплексов и исследования протеома. Нац. Биотехнолог. 17, 7–9 [PubMed] [Google Scholar]
25.
Guerrero C., Tagwerker C., Kaiser P., Huang L. (2006) Интегрированный протеомный подход на основе масс-спектрометрии: количественный анализ тандемных аффинно-очищенных in vivo сшитых белковых комплексов (QTAX) для расшифровки 26 Сеть, взаимодействующая с протеасомами. Мол. Клетка. протеомика
5, 366–378 [PubMed] [Google Scholar]
26.
Kaake RM, Wang X., Huang L. (2010)Профилирование сетей белковых взаимодействий белковых комплексов с использованием аффинной очистки и количественной масс-спектрометрии. Мол. Клетка. протеомика
Мол. Клетка. протеомика
9, 1650–1665 [бесплатная статья PMC] [PubMed] [Google Scholar]
27.
Дайон Л., Хайнард А., Ликер В., Турк Н., Кун К., Хохштрассер Д. Ф., Буркхард П. Р., Санчес Ж.-К. (2008)Относительное количественное определение белков в спинномозговой жидкости человека с помощью МС/МС с использованием 6-плексных изобарических меток. Анальный. хим. 80, 2921–2931 [PubMed] [Google Scholar]
28.
Valli E., Trazzi S., Fuchs C., Erriquez D., Bartesaghi R., Perini G., Ciani E. (2012) CDKL5, новый ген, репрессированный MYCN, блокирует клеточный цикл и способствует дифференцировке нейронных клеток. Биохим. Биофиз. Акта
1819 г., 1173–1185 [бесплатная статья PMC] [PubMed] [Google Scholar]
29.
Сюй П., Дуонг Д.М., Пэн Дж. (2009)Систематическая оптимизация обращенно-фазовой хроматографии для исследовательских статей по дробовой протеомике. Дж. Протеом Рез. 8, 3944–3950 [бесплатная статья PMC] [PubMed] [Google Scholar]
30.
Ван Х., Ян Ю., Ли Ю., Бай Б., Ван С., Тан Х. , Лю Т., Бич Т.Г., Пэн Дж., Ву З. (2015) Систематическая оптимизация масс-спектрометрии хроматографии с длинным градиентом для глубокий анализ протеома головного мозга. Дж. Протеом Рез. 14 829–838 [бесплатная статья PMC] [PubMed] [Google Scholar]
, Лю Т., Бич Т.Г., Пэн Дж., Ву З. (2015) Систематическая оптимизация масс-спектрометрии хроматографии с длинным градиентом для глубокий анализ протеома головного мозга. Дж. Протеом Рез. 14 829–838 [бесплатная статья PMC] [PubMed] [Google Scholar]
31.
Пэн Дж., Элиас Дж. Э., Торин С. С., Ликлайдер Л. Дж., Гиги С. П. (2003)Оценка многомерной хроматографии в сочетании с тандемной масс-спектрометрией (ЖХ/ЖХ-МС/МС) для крупномасштабного анализа белков: дрожжевой протеом. Дж. Протеом Рез. 2, 43–50 [PubMed] [Google Scholar]
32.
Пэн Дж., Шварц Д., Элиас Дж. Э., Торин К. С., Ченг Д., Марсишки Г., Рулофс Дж., Финли Д., Гиги С. П. (2003) Протеомный подход к пониманию убиквитинирования белка. Нац. Биотехнолог. 21, 921–926 [PubMed] [Google Scholar]
33.
Wang X., Li Y., Wu Z., Wang H., Tan H., Peng J. (2014) JUMP: инструмент поиска по базе данных на основе тегов для идентификации пептидов с высокой чувствительностью и точностью. Мол. Клетка. протеомика
13, 3663–3673 [бесплатная статья PMC] [PubMed] [Google Scholar]
34.
Виксон Дж., Келл Д. (2000) Киотская энциклопедия генов и геномов – KEGG. Дрожжи
17, 48–55 [бесплатная статья PMC] [PubMed] [Google Scholar]
35.
Хуан Д.В., Шерман Б.Т., Лемпицки Р.А. (2009 г.) Систематический и комплексный анализ больших списков генов с использованием ресурсов биоинформатики DAVID. Нац. протокол 4, 44–57 [PubMed] [Google Scholar]
36.
Хуанг Д. В., Шерман Б. Т., Лемпицки Р. А. (2009) Инструменты обогащения биоинформатики: пути к всестороннему функциональному анализу больших списков генов. Нуклеиновые Кислоты Res. 37, 1–13 [бесплатная статья PMC] [PubMed] [Google Scholar]
37.
Дженсен Л.Дж., Кун М., Старк М., Чаффрон С., Криви К., Мюллер Дж., Доеркс Т., Жюльен П., Рот А., Симонович М., Борк П., фон Меринг К. (2009 г.) STRING 8 – общий взгляд на белки и их функциональные взаимодействия в 630 организмах. Нуклеиновые Кислоты Res. 37, D412–D416 [бесплатная статья PMC] [PubMed] [Google Scholar]
38.
Бай Б., Чен П., Хейлз С.М., Ву З., Пагала В., Хай А. А., Леви А.И., Лах Дж.Дж., Пэн Дж. (2014) Комплексные подходы к анализу расщепления U1–70K при болезни Альцгеймера. Дж. Протеом Рез. 13, 4526–4534 [бесплатная статья PMC] [PubMed] [Google Scholar]
А., Леви А.И., Лах Дж.Дж., Пэн Дж. (2014) Комплексные подходы к анализу расщепления U1–70K при болезни Альцгеймера. Дж. Протеом Рез. 13, 4526–4534 [бесплатная статья PMC] [PubMed] [Google Scholar]
39.
Терабаяши Т., Фунато Ю., Фукуда М., Мики Х. (2009 г.) Киназа, ассоциированная с везикулами, 104 кДа (CVAK104) индуцирует лизосомную деградацию frizzled 5 (Fzd5). Дж. Биол. хим. 284, 26716–26724 [бесплатная статья PMC] [PubMed] [Google Scholar]
40.
Бриттен Дж. М., Ван Ю., Уилсон С. М., Ханна Р. (2012) Регуляция передачи сигналов CREB через каналы Ca2+ L-типа с помощью Nipsnap-2. каналы
6, 94–102 [PubMed] [Google Scholar]
41.
Хан К.-Дж., Фостер Д.Г., Чжан Н.-Ю., Каниша К., Дзецятковска М., Склафани Р.а, Хансен К.С., Пэн Дж., Лю К.-В. (2012) Убиквитин-специфическая протеаза 9x деубиквитинирует и стабилизирует двигательный нейрон белка выживания спинальной мышечной атрофии. Дж. Биол. хим. 287, 43741–43752 [бесплатная статья PMC] [PubMed] [Google Scholar]
42.
Homan C.C., Kumar R., Nguyen L.S., Haan E., Raymond F.L., Abidi F., Raynaud M., Schwartz C.E., Wood S.a, Gecz J., Jolly L.a. (2014) Мутации в USP9X связаны с Х-сцепленной умственной отсталостью и нарушают миграцию и рост нейронов. Являюсь. Дж. Хам. Жене. 94, 470–478 [бесплатная статья PMC] [PubMed] [Google Scholar]
43.
Li Y., Cheng Y., Huang Y., Conti M., Wilson S.P., Donnell M.O., Zhang H. (2011)Нокаут фосфодиэстеразы-4D и РНК-интерференционный нокдаун улучшают память и усиливают нейрогенез в гиппокампе за счет усиления передачи сигналов цАМФ. Дж. Нейроски. 31, 172–183 [бесплатная статья PMC] [PubMed] [Google Scholar]
44.
Этелл И.М., Паскуале Э.Б. (2005)Молекулярные механизмы развития и ремоделирования дендритных шипов. прог. Нейробиол. 75, 161–205 [PubMed] [Google Scholar]
45.
Стегеман С., Джолли Л. А., Премаратн С., Гец Дж., Ричардс Л. Дж., Маккей-Сим А., Вуд С. а. (2013) Потеря УСП9x нарушает архитектуру коры, развитие гиппокампа и TGFβ-опосредованный аксоногенез. PLoS Один
PLoS Один
8, e68287. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46.
McCrea PD, Gu D. (2010) Краткий обзор семейства катенинов. Дж. Клеточные науки. 123, 637–642 [бесплатная статья PMC] [PubMed] [Google Scholar]
47.
Ван И.-Т. J., Allen M., Goffin D., Zhu X., Fairless A.H., Brodkin E.S., Siegel S.J., Marsh E.D., Blendy J. a, Zhou Z. (2012)Потеря CDKL5 нарушает профиль кинома и связанные с событиями потенциалы, ведущие к аутистическим фенотипам у мышей. проц. Натл. акад. науч. США 109, 21516–21521 [бесплатная статья PMC] [PubMed] [Google Scholar]
48.
Тавьев Ашер Ю.Дж., Скалья Ф. (2012)Молекулярные основы и клинический спектр ранних детских эпилептических энцефалопатий. Евро. Дж. Мед. Жене. 55, 299–306 [PubMed] [Google Scholar]
49.
Чен К., Чжу Ю.-К., Ю Дж., Мяо С., Чжэн Дж., Сюй Л., Чжоу Ю., Ли Д., Чжан С., Тао Дж., Сюн З.-К. (2010) CDKL5, белок, связанный с синдромом Ретта, регулирует морфогенез нейронов посредством передачи сигналов Rac1. Дж. Нейроски. 30, 12777–12786 [бесплатная статья PMC] [PubMed] [Google Scholar]
30, 12777–12786 [бесплатная статья PMC] [PubMed] [Google Scholar]
50.
Рикарди С., Унгаро Ф., Хамброк М., Радемахер Н., Стефанелли Г., Брамбилла Д., Сесса А., Маганьотти К., Бачи А., Джарда Э., Верпелли К., Килструп-Нильсен К., Sala C., Kalscheuer V.M., Broccoli V. (2012) CDKL5 обеспечивает стабильность возбуждающих синапсов за счет усиления взаимодействия NGL-1-PSD95 в постсинаптическом компартменте и нарушается в нейронах, полученных из иПСК пациентов. Нац. Клеточная биол. 14, 1–15 [бесплатная статья PMC] [PubMed] [Google Scholar]
51.
Zhu Y.-C., Li D., Wang L., Lu B., Zheng J., Zhao S.-L., Zeng R., Xiong Z.-Q. (2013) Зависимый от пальмитоилирования CDKL5-PSD-95 взаимодействие регулирует синаптическое нацеливание на CDKL5 и развитие дендритных шипов. проц. Натл. акад. науч. 110, 9118–9123 [бесплатная статья PMC] [PubMed] [Google Scholar]
52.
Тео Г., Лю Г., Чжан Дж., Несвижский А. И., Гинграс А.-К., Чой Х. (2014) SAINTexpress: улучшения и дополнительные функции в анализе значимости программного обеспечения INteractome. Дж. Протеомика
Дж. Протеомика
100, 37–43 [бесплатная статья PMC] [PubMed] [Google Scholar]
53.
Рюдигер А. Х., Рюдигер М., Карл У. Д., Чакраборти Т., Рёпсторфф П., Вехланд Дж. (1999) Подходы на основе аффинной масс-спектрометрии для анализа белок-белковых взаимодействий и сложных смесей пептид-лиганд. Анальный. Биохим. 275, 162–170 [PubMed] [Google Scholar]
54.
Ригетти П.Г. (2014) Король обезьян: личный взгляд на долгий путь к протеомной нирване. Дж. Протеомика
107, 39–49 [PubMed] [Google Scholar]
55.
Сон Р., Ку Б.-К., Юн К.-Дж., Юн М.-Дж., Ю К.-В., Ким Х.-Т., О Х.-Дж., Ким Ю.- Ю., Хан Дж.-К., Ким Ч.-Х., Конг Ю.-Ю. (2006) Neuralized-2 регулирует лиганд Notch в сотрудничестве с Mind bomb-1. Дж. Биол. хим. 281, 36391–36400 [PubMed] [Google Scholar]
56.
Barsi JC, Rajendra R., Wu JI, Artzt K. (2005) Mind bomb1 представляет собой убиквитинлигазу, необходимую для эмбрионального развития мыши и передачи сигналов Notch. мех. Дев. 122, 1106–1117 [PubMed] [Google Scholar]
57.
Юн К.-Дж., Ку Б.-К., Им С.-К., Чон Х.-В., Гим Дж., Квон М.-К., Мун Дж.-С., Мията Т., Конг Ю.-Ю. (2008) Mind bomb 1-экспрессирующие промежуточные предшественники генерируют передачу сигналов Notch для поддержания клеток радиальной глии. Нейрон
58, 519–531 [PubMed] [Google Scholar]
58.
Ямамото М., Морита Р., Мидзогути Т., Мацуо Х., Исода М., Ишитани Т., Читнис А. Б., Мацумото К., Крамп Дж. Г., Ходзуми К., Йонемура С., Каваками К., Ито М. ( 2010) Передача сигналов Mib-Jag1-Notch регулирует формирование паттерна и структурные роли хорды, контролируя решения клеточной судьбы. Разработка
137, 2527–2537 [бесплатная статья PMC] [PubMed] [Google Scholar]
59.
Даскалаки А., Шалаби Н. А., Кукс К., Цумпекос Г., Цибидис Г. Д., Мускавич М. А. Т., Делидакис С. (2011) Различные внутриклеточные мотивы дельты опосредуют ее убиквитилирование и активацию с помощью Mindbomb1 и Neuralized. Дж. Клеточная биология. 195, 1017–1031 [бесплатная статья PMC] [PubMed] [Google Scholar]
60.
Dong Z.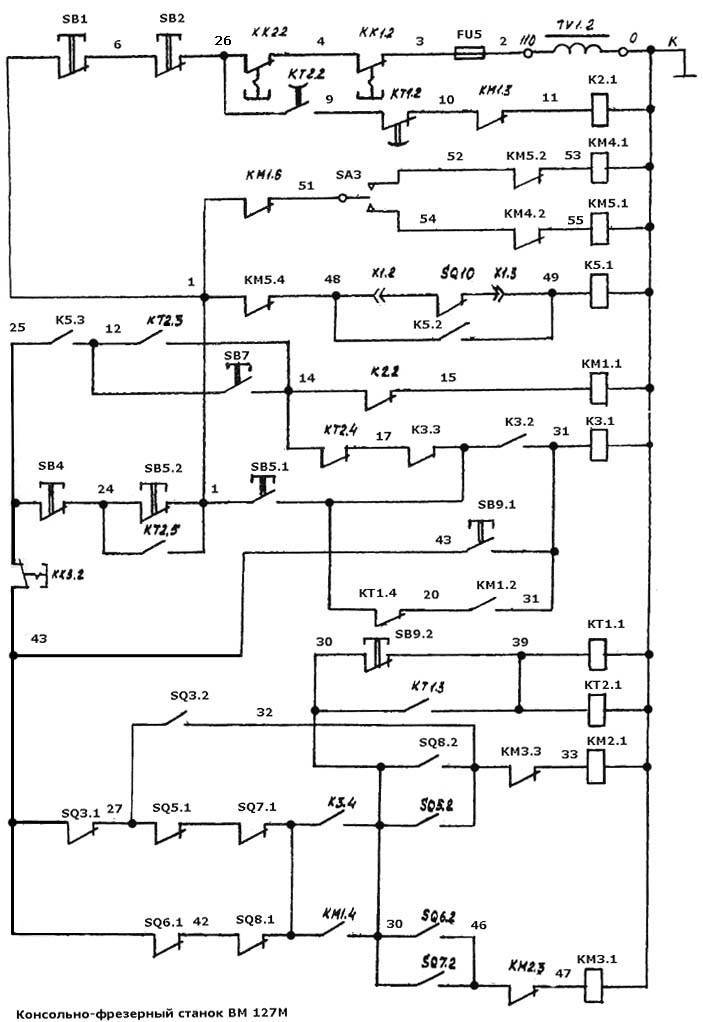 , Yang N., Yeo S.-Y., Chitnis A., Guo S. (2012)Направленная внутрилинейная передача сигналов Notch регулирует самообновление и дифференцировку асимметрично делящейся радиальной глии. Нейрон
, Yang N., Yeo S.-Y., Chitnis A., Guo S. (2012)Направленная внутрилинейная передача сигналов Notch регулирует самообновление и дифференцировку асимметрично делящейся радиальной глии. Нейрон
74, 65–78 [бесплатная статья PMC] [PubMed] [Google Scholar]
61.
Мелоти-Капелла Л., Шергилл Б., Куон Дж., Ботвиник Э., Вайнмастер Г. (2012)Эндоцитоз лиганда Notch создает механическую тяговую силу, зависящую от динамина, эпсинов и актина. Дев. Клетка
22, 1299–1312 [бесплатная статья PMC] [PubMed] [Google Scholar]
62.
Blitzer JT, Nusse R. (2006) Критическая роль эндоцитоза в передаче сигналов Wnt. BMC клеточная биология. 7, 28. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63.
Пак Д. Т., Ян С., Рудольф-Коррейя С., Ким Э., Шэн М. (2001) Регуляция морфологии дендритных шипов с помощью SPAR, RapGAP, связанного с PSD-95. Нейрон
31, 289–303 [PubMed] [Google Scholar]
64.
Боргезиус Н.З., ван Вурден Г.М., Буйтендейк Г.Х.С., Кейзер Н., Яарсма Д., Хоогенраад К.С., Элгерсма Ю.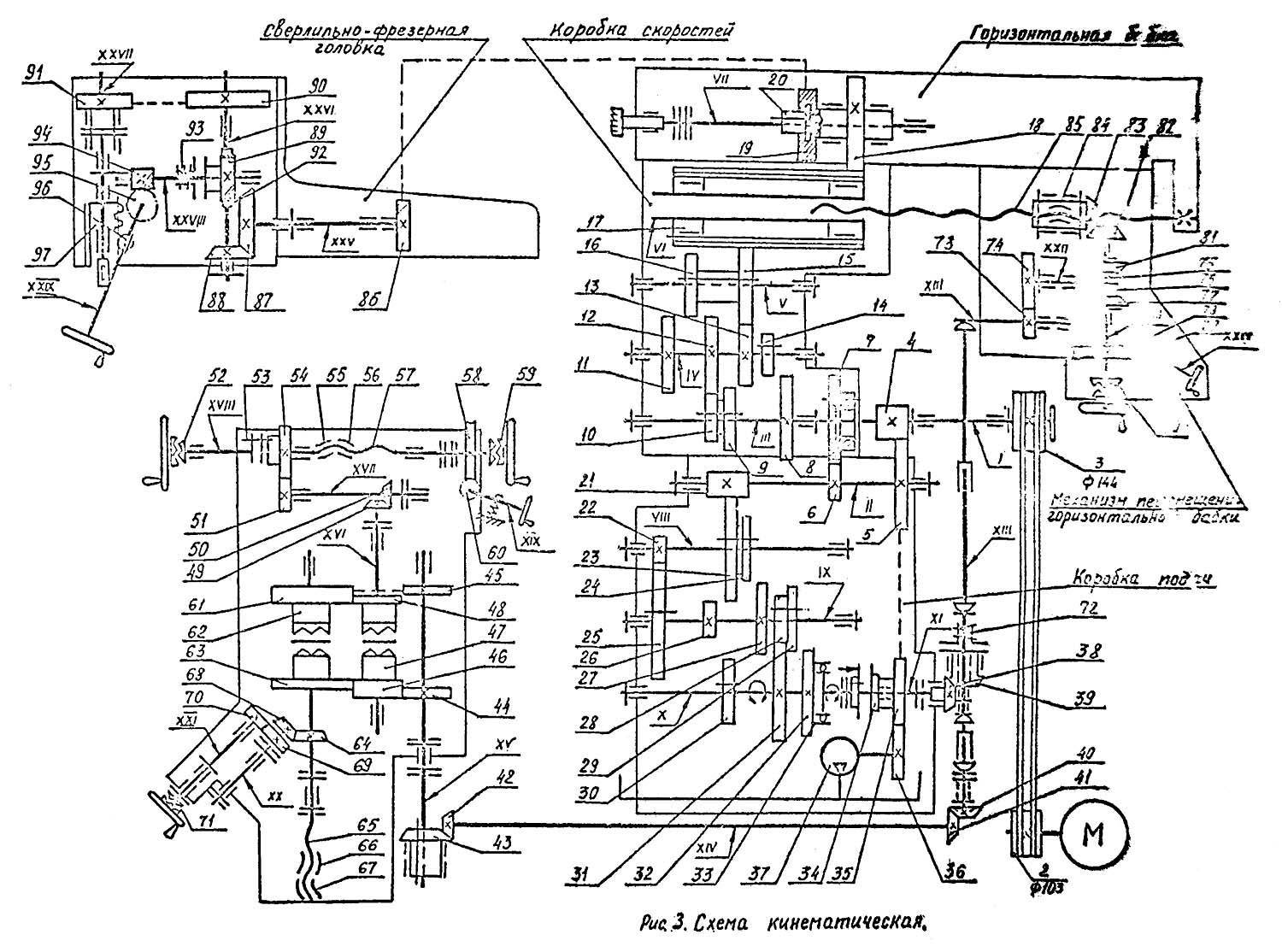 (2011) βCaMKII играет неферментативную роль в синаптической пластичности и обучении гиппокампа, направляя αCaMKII в синапсы. Дж. Нейроски. 31, 10141–10148 [бесплатная статья PMC] [PubMed] [Google Scholar]
(2011) βCaMKII играет неферментативную роль в синаптической пластичности и обучении гиппокампа, направляя αCaMKII в синапсы. Дж. Нейроски. 31, 10141–10148 [бесплатная статья PMC] [PubMed] [Google Scholar]
65.
Südhof TC (2008) Нейролигины и нейрексины связывают синаптическую функцию с когнитивными заболеваниями. Природа
455, 903–911 [бесплатная статья PMC] [PubMed] [Google Scholar]
66.
Fuchs C., Trazzi S., Torricella R., Viggiano R., De Franceschi M., Amendola E., Gross C., Calzà L., Bartesaghi R., Ciani E. (2014) Потеря CDKL5 ухудшает выживаемость и дендритность рост новорожденных нейронов путем изменения передачи сигналов AKT/GSK-3β. Нейробиол. Дис. 70, 53–68 [бесплатная статья PMC] [PubMed] [Google Scholar]
67.
Артузо Р., Менкарелли М. А., Полли Р., Сартори С., Ариани Ф., Поллаццон М., Мароцца А., Чилио М. Р., Спеккио Н., Виджевано Ф., Векки М., Бонивер К., Далла Бернардина Б. , Parmeggiani A., Buoni S., Hayek G., Mari F., Renieri A., Murgia A. (2010)Судорожный вариант синдрома Ретта с ранним началом: определение клинических диагностических критериев.